アラビドプシス体内の鉄ホメオスタシスのための鉄吸収遺伝子発現制御において、FITはAtbHLH38 や AtbHLH39と相互作用している
アラビドプシス体内の鉄ホメオスタシスのための鉄吸収遺伝子発現制御において、FITはAtbHLH38 や AtbHLH39と相互作用している
FIT interacts with AtbHLH38 and AtbHLH39 in regulating iron uptake gene expression for iron homeostasis in Arabidopsis
Youxi Yuan, Huilan Wu1, Ning Wang, Jie Li, Weina Zhao, Juan Du, Daowen Wang, Hong-Qing Ling
Cell Research (2008) 18:385-397. doi: 10.1038/cr.2008.26; published online 12 February 2008
(要約)
鉄は植物の成長と発達に必須の元素である。植物の鉄ホメオスタシスは転写レベルでも転写後レベルでも厳密に制御されている。
近年鉄ホメオスタシスに関わるいくつかのbHLH転写因子群が同定されている。しかしこれらの制御機構は未解明である。本研究ではアラビドプシスにおいて、転写因子FITがAtbHLH38 と AtbHLH39と相互作用し、鉄ホメオスタシスのための鉄吸収遺伝子群の発現制御に直接かかわっていることを示す。
酵母のtwo-hybrid 分析と、アラビドプシスのプロトプラストでのトランジェントアッセイによってAtbHLH38 または AtbHLH39 がアラビドプシスの鉄ホメオスタシスにとって中心的な転写因子であるFITと相互作用をすることを示した。
酵母細胞において、FIT/AtbHLH38 またはFIT/AtbHLH39 による発現は3価鉄キレート還元酵素遺伝子(FRO2)や2価鉄イオントランスポータ遺伝子(IRT1)のプロモーターで駆動させるように設計したGUS遺伝子の発現を活性化させた。
植物体内でFIT遺伝子をAtbHLH38遺伝子またはAtbHLH39遺伝子と共に過剰発現させた場合には、FRO2遺伝子 とIRT1遺伝子が誘導的発現から恒常的発現に転換した。
さらにこれらの過剰発現植物では、鉄投与の有無にかかわらず、IRT1タンパクの集積や高い3価鉄還元活性が検出されたので、FRO2遺伝子 とIRT1遺伝子は転写後制御されているのではないことがわかった。
異なる遺伝子を二重に組み合わせた過剰発現植物は、野生型や、単にAtbHLH38, AtbHLH39 , FITを過剰発現させたものよりも多くの鉄を地上部に集積した.
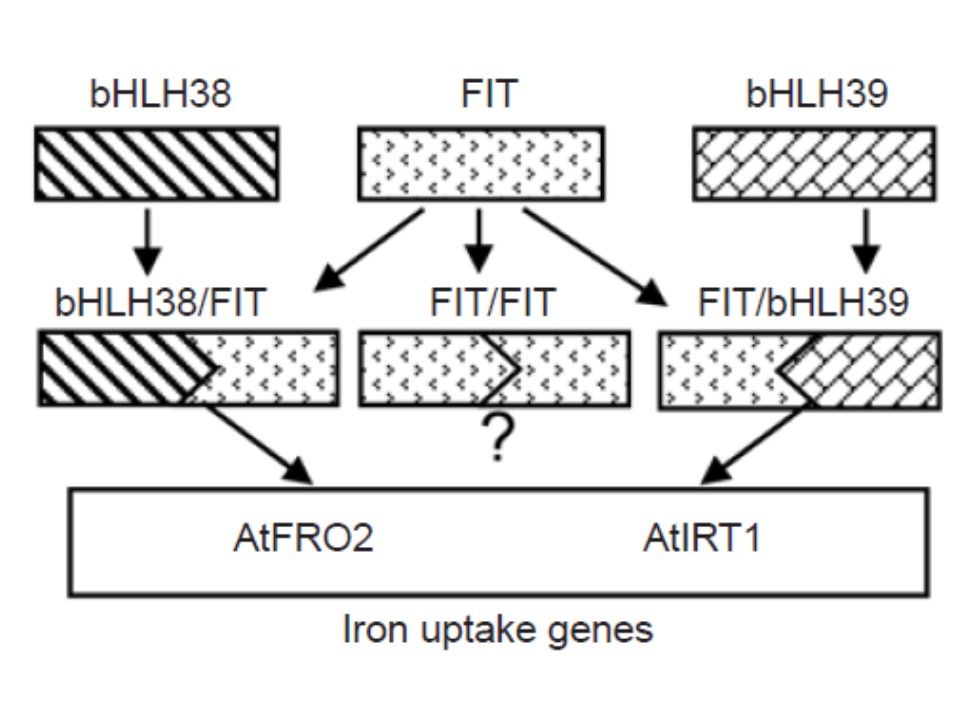
我々のデータから言えることは、FRO2遺伝子とIRT1遺伝子がこれら3つの転写因子のターゲットであり、FRO2遺伝子とIRT1遺伝子はFIT/AtbHLH38 またはFIT/AtbHLH39という複合タンパクによって制御されているということである。
(下図の説明)
FRO2遺伝子とIRT1遺伝子に働く転写因子FIT, AtbHLH38および AtbHLH39の制御モデル。FITは鉄欠乏ストレス条件下でのアラビドプシスで、効果的な鉄吸収を促進するのに必須の転写因子である。FRO2遺伝子とIRT1遺伝子の発現の活性化には、AtbHLH38 か AtbHLH39タンパクがFITタンパクとヘテロダイマーを形成することが必要である。