僶僀僆僀儞僼僅乗儅僥僀僢僋僗偺庤朄傪梡偄偨僀僱偺揝夁忚墳摎婡峔偵娭學偡傞怴婯偺僔僗惂屼攝楍偲僾儘儌乕僞乕峔憿偺夝柧
埲壓偺榑暥偼丄揝夁忚偵墳摎偡傞怴婯僔僗惂屼攝楍偺敪尒偱偁傞丅猕孨傗傝傑偟偨偹丅偍傔偱偲偆両
丂丂
丂丂丂
僶僀僆僀儞僼僅乗儅僥僀僢僋僗偺庤朄傪梡偄偨僀僱偺揝夁忚墳摎婡峔偵娭學偡傞怴婯偺僔僗惂屼攝楍偲僾儘儌乕僞乕峔憿偺夝柧
Elucidation of Novel cis-Regulatory Elements and Promoter Structures Involved in Iron Excess Response Mechanisms in Rice Using a Bioinformatics Approach
丂丂
Yusuke Kakei, Hiroshi Masuda, Naoko K. Nishizawa, Hiroyuki Hattori and May Sann Aung
丂丂丂
乮梫栺乯
揝乮Fe乯夁忚偼扺悈忦審壓偺巁惈搚忞偱偼丄嶌暔惗嶻丄摿偵僀僱偺嵧攟偱偺庡梫側惂栺忦審偲側偭偰偄傞丅
丂
揝夁忚壓偱偼怉暔偼暋嶨側儊僇僯僘儉偲丄崻偐傜偺揝偺攔弌傗條乆側慻怐偱揝偺妘棧傪峴偆僱僢僩儚乕僋傪丄妶惈壔偝偣傞丅
丂
僀僱偱偼揝夁忚墳摎惂屼偺儊僇僯僘儉偱偁傞揮幨場巕傗僔僗惂屼攝楍(CREs)側偳偼傑偩傑偩晄柧偱偁傞丅
丂
変乆偼埲慜偵條乆側僀僱偺慻怐偱條乆側儗儀儖偱偺揝夁忚偵墳摎偡傞曪妵揑側儅僀僋儘傾儗僀夝愅偵偮偄偰曬崘偟偨丅
丂
杮曬崘偱偼丄偙偺儅僀僋儘傾儗僀夝愅僨乕僞傪梡偄偰丄僀僱偺怴偟偄CRE倱偲僾儘儌乕僞乕峔憿傪丄僶僀僆僀儞僼僅儅僥僀僢僋僗偺庤朄傪梡偄偰偝傜偵奐柧偟偨丅
丂
傑偢変乆偼揝夁忚娭楢CREs傪梊應偡傞偨傔偵揝夁忚墳摎揮幨reguron傪僇僥僑儔僀僘偟偰丄4偮偺庡梫側僋儔僗僞乕傪尒弌偟偨丗揝挋憼宆丄揝僉儗乕僩宆丄揝媧廂宆丄偦傟偵WRKY偲偦偺懠偺嫟敪尰僞僀僾偱偁傞丅
丂
師偵変乆偑偙傟傜4偮偺堚揱巕敪尰僋儔僗僞乕偺側偐偺CREs傪扵嶕偟偨偺偼丄変乆偑埲慜偵奐敪偟偰偄偨microarray-associated motif analyzer (MAMA)偲屇偽傟傞庤朄傪梡偄偰偱偁傞丅
丂
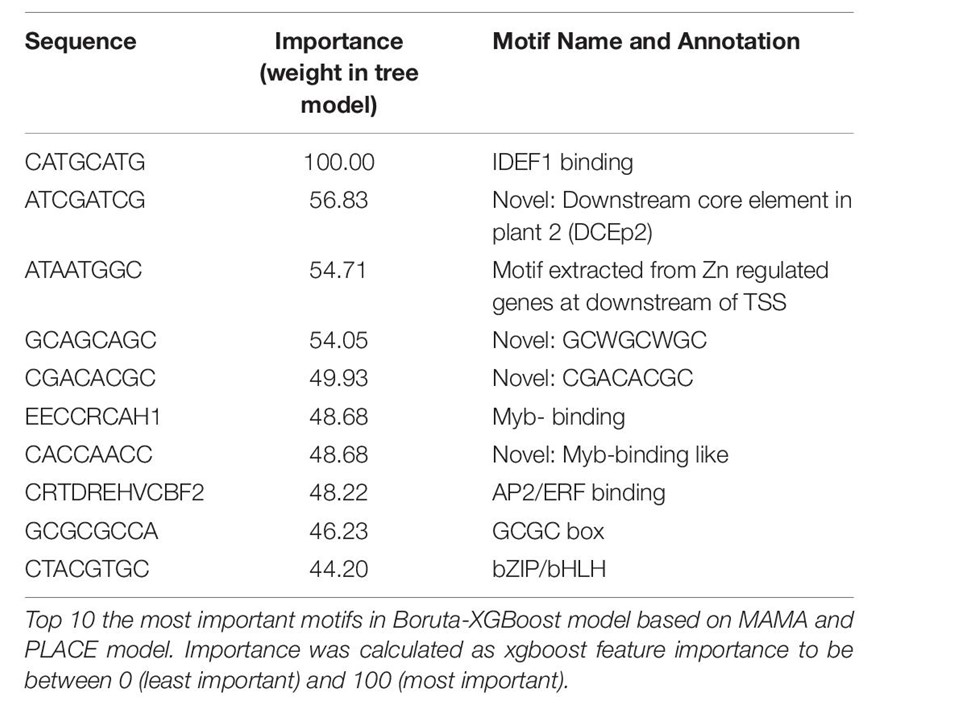
曪妵揑側僶僀僆僀儞僼僅乕儅僥僀僋僗傪捠偟偰丄変乆偼MAMA暘愅偵傛偭偰慡晹偱俆俇侽屄偺CRE傪拪弌偟丄CRE倱偺偆偪偺條乆側僀僱偺慻怐偱揝夁忚墳摎偵捈愙娭學偡傞42屄偺廳梫側曐懚攝楍傪拪弌偟偨丅
丂
変乆偼揝夁忚CREs偺怴婯偺cis-攝楍岓曗偲偟偰GCWGCWGC, CGACACGC, 偦偟偰 Myb寢崌條儌僠僀乕僼傪扵偟摉偰偨.
丂
MAMA傗傛偔抦傜傟偨PLACE CREs偺庤朄傪梡偄偨岓曗CREs偺桳柍傪婎偵丄Boruta-XGBoost儌僨儖偱偺敪尰僷僞乕儞傪栺83亾偲偄偆崅偄惛搙偱愢柧偱偒傞偙偲傪尒弌偟偨丅
丂
怴婯偺MAMA CREs偲婛抦偺PLACE CREs偵傛偭偰擹弅偝傟偨攝楍偼崅偄惛搙偺敪尰僷僞乕儞傪帵偟偨丅
丂
変乆偼傑偨揝夁忚墳摎偺婛抦偺CREs偺怴偟偄栶妱傕尒弌偟偨丄椺偊偽DCEp2 motif, IDEF1-, Zinc Finger-, WRKY-, Myb-, AP2/ERF-, MADS- box-, bZIP 偲丄揝夁忚墳摎堚揱巕孮偺拞偺 bHLH-寢崌攝楍
丂
偝傜偵怴婯偵敪尒偝傟偨CRE倱偵傕偲偯偄偰揝夁忚墳摎堚揱巕孮偺暘巕儌僨儖偲僾儘儌乕僞乕峔憿傪峔抸偟偨丅
丂
憤妵偡傞偲揝夁忚娭楢CREs偲曐懚攝楍偵娭偡傞変乆偺敪尒偼丄揝夁忚墳摎戙幱偵娭學偡傞堚揱巕傗揮幨場巕偺敪尒傗丄僀僱偺揝夁忚墳摎偺夝柧傗丄彨棃偼僾儘儌乕僞乕攝楍偺揝夁忚懴惈昳庬偺嶌弌傊偺墳梡側偳丄憤妵揑側帒尮傪採嫙偡傞偙偲偵側傞偩傠偆丅
丂
丂
丂
埲壓昞偲恾偺愢柧
丂
昞1丂堚揱巕敪尰僷僞乕儞儌僨儖偺廳梫側儌僠乕僼
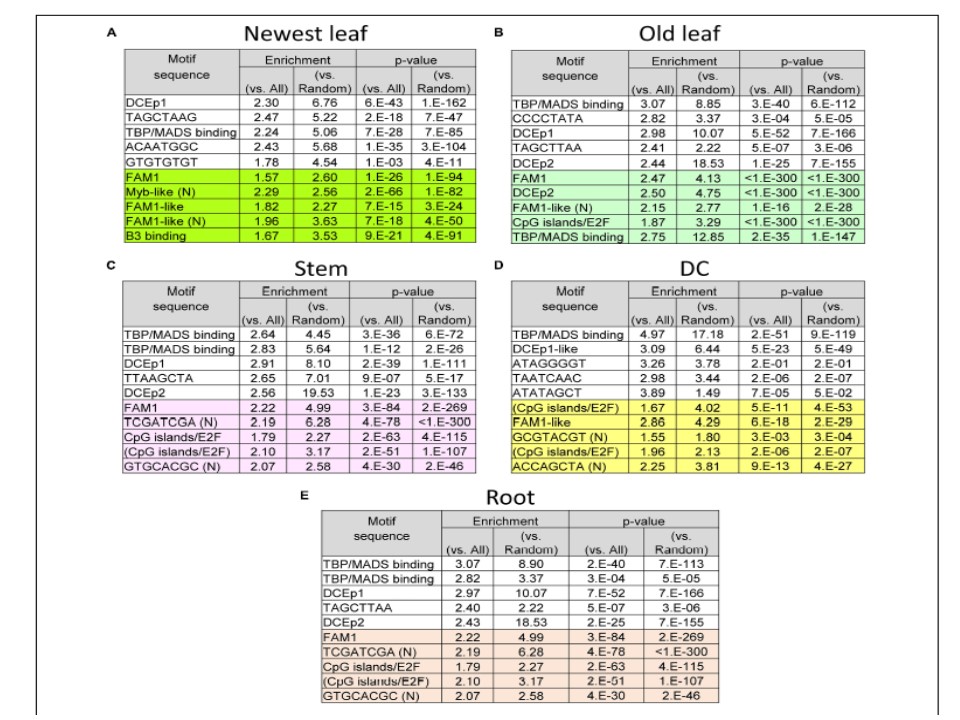
昞2丂奺慻怐偐傜拪弌偝傟偨CRE岓曗
乮A乯嵟怴梩丄乮B乯屆偄梩丄乮C乯宻丄乮D乯DC丄乮E乯崻丅DC偼discrimination center偺棯偱丄崻偲梩偺寢崌愡埵丅
丂丂
敀偄攚宨偺僙儖偼僂僀儞僪僂暆–50偐傜+150娫偺CRE岓曗丅怓晅偒偺攚宨偼TSS偺俆侽侽倐倫忋棳偺C俼俤岓曗丅乮俶乯偼偙傟傑偱偵曬崘偑側偐偭偨俠俼俤丏
丂丂
CpG islands/E2F (CGCGCGCA)儌僠乕僼偼CGCGCGCG, CGCGCGTA, CGCGCGTG偺堎宍偱丄乬(CpG islands/E2F)乭偲拲庍偝傟偰偄傞丅
丂
儌僠乕僼偺棙摼偼丄(number in regulon)/(total length of 500 bp upstream or –500 bp to +150 bp sequence of regulon)傪(number in all genes)/(total length of upstream 500 bp sequence of all genes)偱妱偭偨傕偺偱偁傞丅
丂丂
P-抣偼number of motifs found in regulon, number of regulon genes, chance of motif found in all genes側偳傪巊偆擇崁幃僥僗僩偱寁嶼偟偨丅
棙摼偲p亅抣傕摨偠偔丄偡傋偰偺堚揱巕攝楍偺偐傢傝偵儔儞僟儉攝楍偺摨偠僒僀僘偺傕偺傪梡偄偰寁嶼偟偨丅
丂丂
丂丂丂
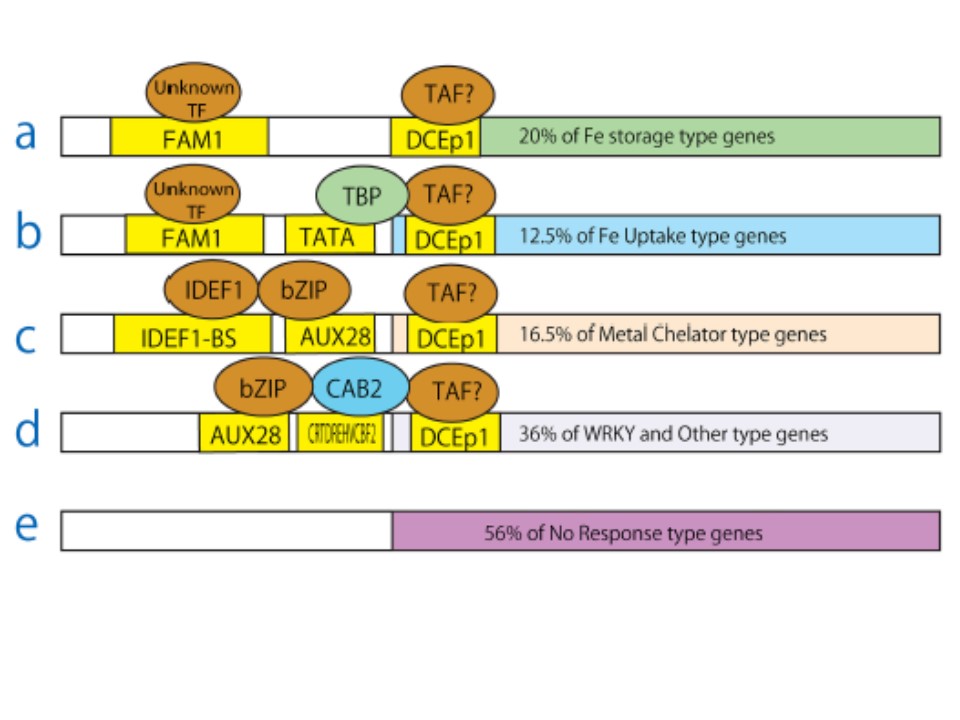
恾侾丂揝夁忚墳摎堚揱巕惂屼偺戙昞揑側暘巕儌僨儖
揮幨場巕IDEF1偼CATGCATG偵寢崌偟丄bZIP偼SGBFGMGMAUX28偵寢崌偟丄TBP偼CTATAAAT偵寢崌偟丄CAB2偼CRTDREHVCBF2偵寢崌偡傞丅
丂丂
CTACACCT, FAM1, and DCEp1偵寢崌偡傞揮幨場巕(TFs)偼枹抦偱丄TAF, TBP associated factor.偲偟偰乬Unknown TF乭偲帵偟偰偁傞丅
亾偼埲壓偺傛偆偵寁嶼偟偨((Percentage of specific type genes in specific branch) 亊 (percentage of all types in specific branch))/(percentage of specific type (20%)/percentage of all types (100%)).丂椺偊偽丄B儌僨儖偵婰嵹偝傟偰偄傞12.5%偺揝媧廂僞僀僾堚揱巕 攝壓偺傛偆偵寁嶼偝傟偨丗(0.05 * 0.50)/(20/100) = 0.125.
昞1
昞2
恾1