DNAのメチル化はイネ(Oryza sativa)の鉄欠乏順化に関係している
DNAのメチル化はイネ(Oryza sativa)の鉄欠乏順化に関係している
DNA methylation is involved in acclimation to iron-deficiency in rice (Oryza sativa)
Shuo Sun, Jiamei Zhu, Runze Guo, James Whelan and Huixia Shou
The Plant Journal (2021) doi: 10.1111/tpj.15318
Received 24 February 2021; revised 27 April 2021; accepted 3 May 2021. *For correspondence (e-mail: huixia@zju.edu.cn).
要約
鉄(Fe)は必須微量元素であり、鉄を制限すると生育、収量、食品としての質に有意に影響を及ぼす。
鉄制限に関する応答に関しては転写プロファイルが報告されており、分子メカニズムの研究が遂行されているが、鉄欠乏に応答するエピジェネテイック因子の役割に関してはほとんどわかっていない。
本研究では、イネ幼植物を用いて鉄十分条件下と鉄欠乏条件下で、根部と地上部について、 全ゲノム bisulfite シーケンス分析, high-throughput RNA-Seq of mRNA, small RNA、 転移因子(TE)の発現について調べた。
その結果鉄欠乏処理によって、広範囲に、特にCHH配列において、異常なメチル化が起こっていることが明らかになった。(:HはC以外の核酸塩基)
メチル化と転写因子群を統合的に分析すると、鉄欠乏誘導性遺伝子の転写量は、転移因子群TEs)近傍でネガテイブに相関があり、24-nucleotide siRNAsとはポジテイブに相関があった。
鉄欠乏応答に対するメチル化の生理学的分子生物学的可能性を、DNAメチルトランスフェラーゼ阻害剤(5-azacytidine)を外部投与する方法と、ドメインを転位させたメチルトランスフェラーゼ2(DRM2)のCHHメチル化欠損変異株を用いて検定した。
両方のアプローチともイネの生育と鉄含量を低下させた。
したがって、特異的なメチル化パターンが siRNAsの指示によって変化させられるということが鉄欠乏条件に対する順化にとって重要な役割を担っていることがわかった。
さらに、(Piなどの)ほかの論文と比較すると、この事は栄養を制限された条件に対する共通の順化メカニズムであるのかもしれない。
緒言
(以下は部分訳です:メチル化のメカニズムは複雑なので、一部文献も示しています)
シトシンDNAメチル化(mC)は、動植物におけるゲノムの安定性と遺伝子発現における主要な役割を演じるエピジェネテイックな特徴である。
(He and Ecker, 2015; Huff and Zilberman, 2014; Schmitz et al., 2019; Zhang et al., 2018).
それは転移因子(TEs)の転写のサイレンシング(静止)に関係して、ゲノムの正確性の維持に貢献すると同時に、特定の遺伝子発現制御にも関係している。
(Law and Jacobsen, 2010; Rodrigues and Zilberman, 2015).
シトシンのメチル化は、CG 、 CHG (対称的) 、 CHH (非対称的) (; ここで H = C, AまたはT )の3つの配列で進行する。
それぞれの配列のメチル化は特異的DNAメチルトランスフェラーゼによって活性化される。
(Law and Jacobsen, 2010).
CG と CHGの修飾はDNA複製時には半保存的にDNAメチルトランスフェラーゼ(MET1)(Kankel et al., 2003)によって制御されている。またchromomethylase 3 (CMT3)はH3K9メチル化を認識することによってDNAを標的としている。(Bartee et al., 2001; Lindroth et al., 2001).
新規のGHHメチル化はDNA複製後に生じ、RNA-directed DNA メチル化(RdDM)回路
と相互作用しながらsmall(nts) [典型的な24塩基]によって確立する。
(Mirouze and Paszkowski, 2011).
RdDMにおいては、RNAポリメラーゼ IVと Dicer-様タンパクがsiRNA 生合成に必要だが、RNAポリメラーゼIV、AGO (argonaute) 4/6、メチルトランスフェラーゼドメイン再構成 メチルトランスフェラーゼ 2 (DRM2)が、siRNA-によってガイドされたDNAメチル化に関係している。
(Kawashima and Berger, 2014; Law and Jacobsen, 2010; Matzke et al., 2015; Matzke and Mosher, 2014; Nuthikattu et al., 2013).
DNAメエチル化の状態はDNAメチルトランスフェラーゼばかりでなく、いくつかのDNA脱メチル化酵素、たとえばROS1 (repressor of silencing 1)、DME (Demeter), DML2 (Demeter-like 2)、 DML3によっても維持されている。
(Liu et al., 2018; Ortega-Galisteo et al., 2008; Penterman et al., 2007).
植物が、温度、光強度、栄養、水の利用性などでの不利な環境に曝されると、植物はそれらの不良事象をDNAメエチル化パターン変化で「記録」する。
(Kinoshita and Seki, 2014).
無機リン(Pi)が制限条件下で生育したイネに再度リンを投与した場合のmCのspatio-temporal dynamics(時空ダイナミックス)に関する研究では、Pi欠乏誘導性変化mCのなかでは、CHHコンテクストが転写産物が増加していた約205 bp遺伝子近傍の転写因子(TEs)に優先的に局在していた。
(Secco et al., 2015).
さらに、これらのmC変化は近傍遺伝子の転写産物の増加後も起こっているので、有糸分裂を通じて部分的に遺伝されるのかもしれない。
さらに、これらのメチル化パターンの変化は、通常のRdDM回路に関係しているキーファクターであるDCL3a (Dicer-like 3a)とはおおむね独立している。
他の研究によると、CG メチル化とCGの非メチル化の広範囲にわたる違いが、カドミウム(Cd)に曝されるか、Cdに曝されていないかのイネゲノムに記録されていることが同定されている。
(Feng et al., 2016).
野生型に比べるとメチル化の維持に必要なMET1 と DRM2の変異株は、Cdストレス下での金属輸送体遺伝子、Cd解毒タンパク、金属関連転写因子(TFs)の転写レベルを低下させる。
包括的なDNAメチル化阻害剤である5-azacytidineはCd応答性遺伝子のDNAメチル化レベルを全般的に減少させるが、イネ幼植物の生長や、体内Cd集積は増加した。
2種類のPi利用効率の異なるダイズの品種でのDNAメチル化地図を比べると2つの品種ともにPi制限条件下でメチル化の程度がわずかに高かったが、Pi欠乏下でsiRNAの含量の変化に応答して、CHHメチル化配列での転移因子(TE)の領域でダイナミックなメチル化の変化が起こっている。
(Chu et al., 2020).
亜鉛(Zn)に関しては、アラビドプシスの根のZn欠乏処理を長く続けると、ある種のZn欠乏誘導性遺伝子のアップレギュレーションに関連して、CG と CHGの配列間で異なるメチル化が起こるがCHH配列では起こらなかった。
(Chen et al., 2018).
本論文では、鉄欠乏と鉄十分条件下でのイネの地上部と地下部において、DNAメチローム、タンパクをコードする遺伝子のトランスクプリトーム、転移因子(TEs)、siRNAプロファイルを決定した。
鉄欠乏に応答して特にCHHコンテキストでの広範囲な高メチル化(ハイパーメチレーション)領域が観察された。鉄欠乏条件下での転写量の変化に関して、siRNA 変化以外にはDNA methyltransferase やその他の RdDM 成分等には有意な変化が見られなかった。
本研究ではイネの鉄欠乏条件下でのDNAメチル化の動態を明らかにし、栄養欠乏条件下での応答として、可動性siRNA依存性RdDM (mobile siRNA-dependent RdDM mechanism) のメカニズムがあることを示唆した。
実験方法
(略)
論議
(部分訳です)
過去10年間に様々なトランスクリプトーム解析や機能解析の研究で、植物がどのようにして鉄ホメオスタシスを維持しているかに関しての知見が大きく集積してきた。
(Kobayashi and Nishizawa, 2012; Kobayashi et al., 2019).
しかし鉄欠乏応答におけるエピゲノム変化の役割の可能性についてはわずかしかわかっていない。
本研究で、我々は鉄十分条件下と鉄欠乏条件下でのイネの根部と地上部における、DNAメチル化一塩基分解能地図(single base resolution map of DNA methylation)を作成した。
鉄十分条件下でも鉄欠乏条件下でもゲノムのシトシン(C)メチル化レベルは似ているのだが、鉄欠乏で全体的にDNAメチル化レベルが増加したCHH配列部位では特異的なメチル化の変動が観察された。
さらに、methyltransferase阻害剤であるAza(5-azacytidine)でイネを処理すると鉄シグナル経路遺伝子群が抑制されて、鉄欠乏下でのイネの鉄含量が低下した。
次に、鉄欠乏応答へのメチル化の役割を確認するために遺伝学的アプローチを行った。
osdrm2変異体は野生株と比較して全ゲノムレベルでCHHメチル化レベルが有意に低下しており、生育阻害とSPAD値の低下が、鉄十分条件下と鉄欠乏条件下で、ともに観察された。
それほど多くの鉄欠乏誘導性遺伝子が鉄欠乏条件下で高メチル化(hypermethylation)しているわけではないが、鉄欠乏応答転写因子であるIRO2 と OsbHLH156が高メチル化していた。
以上を総合して、我々はIRO2 とOsbHLH156のメチル化変化は、これらの遺伝子が多くの鉄欠乏応答の下流の標的遺伝子の誘導に必要であるので、重要な制御特性であると提案したい。
これと同様の例が以前にイネのリン酸欠乏でも見いだされている。
(Secco et al., 2015).
その研究では、経時的にデータ解析しており、転写がDNAメチル化の以前に起こっていると提唱されている。
であるから、遺伝子の転写誘導によってDNAメチル化が引起こされると考えられる。
さらに、DNAメチル化は鉄欠乏とリン欠乏で有意差がない。しかも、鉄欠誘導性とリン欠誘導性遺伝子に関してメチル化領域は、共にCHH配列と転移因子(TE)に及んでいる。
これらの特質は環境中での栄養制限条件に応答するメカニズムが類似していることを示唆している。
この仮説はリン欠乏やその他の栄養欠乏条件下でのsmall RNA発現プロファイルをもちいてさらに確認する必要がある。
24-nt siRNAsの集積はRdDM(RNA-directed DNA メチル化)回路 活性の重要な指標となるものである。
(Law and Jacobsen, 2010; Matzke et al., 2015)
本件研究の結果では、鉄欠乏誘導性遺伝子群に隣接する転移因子(TE)領域に多数の 24-nt siRNAsが補充され、転移因子群(TEs)の転写量が鉄欠乏で抑制された。
一方 高CHHメチル化レベルが24-nt siRNA領域と転移因子(TE)本体で観察されたことは、 RdDM(RNA-directed DNA メチル化) 活性が、鉄欠乏応答下でのメチル化において起こる特異的変化のもとに横たわるメカニズムによるかもしれない事を示唆している。
しかし、RDR1-6、AGO4/6 、 DCL3など24-nt siRNAsの生成に関わる遺伝子をコードする遺伝子の転写産物量は、鉄欠乏下でも変化がなかった。
本研究では「23-nt と24-nt 高 siRNA クラスター」が地上部よりも根部で多く検出されたが、「低 siRNAクラスター」が根部よりも地上部で多く検出された。
これまでの多くの研究では、23–24-nt siRNAsが地上部と根部の間を移動する主な長距離可動性siRNAsであることが明らかになっている。
(Martınez et al., 2016; Molnar et al., 2010; Pagliarani and Gambino, 2019; Tamiru et al., 2018).
であるから、遺伝子発現を変動させるためのsiRNAのクラスター量の変化は、地上部と根部の間のsiRNAの長距離輸送に依存している。
以上を総合するとイネの鉄欠乏への順化はゲノム中の特異的遺伝子に隣接するCHHメチル化の増加によるものであると考えられる。
このような局所的な超メチル化(hypermethylation)は通常のRdDM(RNA-directed DNA メチル化) -関連酵素をコードする遺伝子の転写産物量の変化に依存するものではなく、根部と地上部間での可動性siRNAsの移動に依存するものであると考えられる。
24-nt siRNAが転移因子(TEs)にリクルートされてそのDNAメエチル化のレベルを上げて、隣接遺伝子、特に鉄欠乏応答経路のキーとなる調節遺伝子の遺伝子発現に対して負の効果を及ぼしているTEsを阻害するのである。
このようなメカニズムで鉄欠乏遺伝子誘導が増強されてそれが長期間維持されて、鉄欠乏条件下での鉄吸収が最適化される。
(この鉄に関する結果を)メチル化とトランスクリプトームにおけるPi欠乏誘導性変化の報文と併せて検討すると、これがイネの栄養欠乏ストレスへの適応の普遍的メカニズムと考察される。
これらの発見は非生物的ストレスに対する植物の応答における、DNAメチル化と小分子RNAの役割に対する知見を広めるものである。
図1の説明
茎葉の鉄欠乏シグナル経路の上流におけるコアな転写因子(TFs)であるIRO2とOsbHLH156の変化をlog2[-Fe/+Fe]で示している。
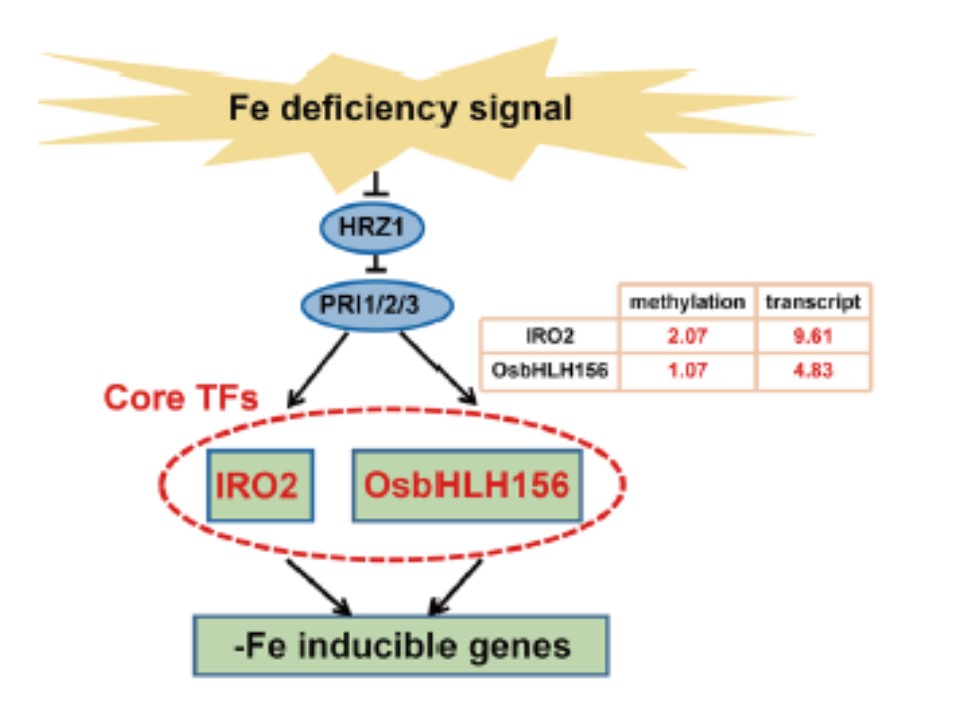
図1