憤愢丗塰梴慺偺儂儊僆僗僞僔僗傪挷愡偡傞儅僀僋儘RNA丗僶僀僆僼僅乕僥傿僼傽僀僪嶌暔奐敪偺偨傔偺帩懕壜擻側傾僾儘乕僠
嵟嬤丄塰梴慺偺戙幱傗桝憲偵娭學偡傞堚揱巕偺敪尰偵娭傢傞丄條乆側儅僀僋儘傾儗僀偺尋媶惉壥偑懕乆偲敪尒偝傟偰偄傞丅拞偱傕拏慺偵娭偡傞儅僀僋儘傾儗僀偑8偮偱丄揝偵娭偟偰傕俉偮偁傝丄忋埵傪愯傔偰偄傞丅偙偙偱偼丄揝偵娭偡傞晹暘偩偗徯夘偡傞丅偙偺榑暥偺堷梡暥專棑偱尒傞尷傝丄側偤偐偙偺曽柺偺擔杮偺尋媶幰偼旕忢偵彮側偄傛偆偱偁傞丅
丂丂
丂丂
憤愢丗塰梴慺偺儂儊僆僗僞僔僗傪挷愡偡傞儅僀僋儘RNA丗僶僀僆僼僅乕僥傿僼傽僀僪嶌暔奐敪偺偨傔偺帩懕壜擻側傾僾儘乕僠
丂
MicroRNAs modulating nutrient homeostasis: a sustainable approach
for developing biofortified crops
丂
Monica Jamla丒 Shrushti Joshi丒 Suraj Patil丒 Bhumi Nath Tripathi丒 Vinay Kumar
奣梫
柍宻惈怉暔偼丄偦偺庻柦偺娫丄嵟揔偱側偄塰梴墫擹搙偺惗暔妛揑棙梡擻偵懳張偟丄揔愗側惉挿偲戙幱偺偨傔偵丄岠棪揑側塰梴愛庢丄挋憼丄堏摦偺偨傔偺僔僌僫儖僇僗働乕僪偺楢寢栐傪忢偵姶抦偟恑壔偝偣側偗傟偽側傜側偄丅
偟偐偟丄娐嫬偺曄摦偲恖堊揑側妶摦偺寖壔偼丄怉暔偵偲偭偰庤偛傢偄壽戣偲側偭偰偄傞丅
偙偺偙偲偑丄乮旝乯塰梴晄懌偺嶌暔傗塰梴晄埨偵攺幵傪偐偗偰偄傞丅僶僀僆僼僅乕僥傿僼傿働乕僔儑儞偼丄旝検塰梴慺偺塰梴幐挷偲愴偆偨傔偵棙梡偱偒傞丄帩懕壜擻偱岠壥揑側傾僾儘乕僠偲偟偰拲栚偝傟偰偄傞丅
僶僀僆僼僅乕僩壔嶌暔偼丄廬棃偺堢庬丄擾妛揑庤朄丄傑偨偼愭恑揑側僶僀僆僥僋僲儘僕乕僣乕儖傪梡偄偰奐敪偝傟偨丄朷傑偟偄塰梴慺偺儗儀儖偑嫮壔偝傟偨嶌暔偱偁傞丅傑偨偼崅搙側僶僀僆僥僋僲儘僕乕傪嬱巊偟偰奐敪偝傟偨傕偺偱偁傞丅
塰梴慺偺儂儊僆僗僞僔僗偼丄塰梴慺僗僩儗僗壓偱慾奞偝傟丄暋悢偺嵶朎偍傛傃暘巕峔惉梫慺偑娭梌偡傞抁嫍棧偍傛傃挿嫍棧偺嵶朎-嵶朎/憻婍娫偺僐儈儏僯働乕僔儑儞偵忈奞偑敪惗偡傞丅
僶僀僆僀儞僼僅儅僥傿僋僗僷僀僾儔僀儞傗僨乕僞儀乕僗偲楢摦偟偨崅搙側僔乕働儞僒乕偵傛傝丄彫偝側僔僌僫儖揱払暘巕傗揮幨屻惂屼場巕偱偁傞儅僀僋儘RNA乮miRNA乯偑丄塰梴峆忢惈傪娷傓怉暔偺庡梫尰徾偵娭梌偟偰偄傞壜擻惈偑帵嵈偝傟偰偄傞丅
miRNA偼丄僶僀僆僥僋僲儘僕乕偵婎偯偔惗暔嫮壔僾儘僌儔儉偺怴偨側僞乕僎僢僩偲偟偰拲栚偝傟偰偄傞丅
偙偺傛偆偵丄miRNA偺婡峔揑摯嶡偲惂屼揑栶妱傪棟夝偡傞偙偲偼丄塰梴岠棪偺崅偄僶僀僆僼僅乕僩嶌暔偺奐敪偵偍偄偰丄偦傟傜傪扵媮偡傞偨傔偺怴偟偄憢傪奐偔壜擻惈偑偁傞丅
杮憤愢偱偼丄怉暔塰梴妛偲塰梴峆忢惈偵偍偗傞miRNA偺堄媊偲栶妱偵偮偄偰弎傋傞丅
傑偨丄庡梫塰梴慺偱偁傞拏慺丄儕儞丄棸墿丄儅僌僱僔僂儉丄揝丄垷墧偵懳偡傞怉暔偺斀墳偵廳梫側栶妱傪壥偨偡偙偲傕夝愢偟偰偄傞丅
miRNA傪慻傒崬傫偩僶僀僆僼僅乕僩嶌暔偺奐敪偵偮偄偰丄嵟嬤偺惉岟帠椺傪岎偊偰揥朷偟偨丅傑偨丄尰嵼偺壽戣偲彨棃偺愴棯偵偮偄偰傕弎傋偨丅
丂
埲壓奺塰梴慺偵懳墳偡傞儅僀僋儘RNA偺夝愢偑偁傞偑丄偙偙偱偼偦偺偆偪偺揘偵捛偆偲偡傞儅僀僋儘傾儗僀偵娭偟偰偺崁栚偺傒栿弌偟偨丅
丂丂
揝偵墳摎偡傞儅僀僋儘RNA
揝乮Fe乯偼搚忞拞偵尷掕揑偵傑偽傜偵暘晍偟丄拞惈傑偨偼墫婎惈搚忞偱偼偦偺梟夝搙偐傜棙梡偱偒側偄偙偲偑懡偄偙偲偑娤嶡偝傟偰偄傑偡丅巁惈搚忞偱偼丄寵婥惈忦審偵傛傝揝偺撆惈偑桿敪偝傟傞(Hell and Stephan 2003)丅揝寚朢壓偱偼丄揝寚朢墳摎惈僔僗嶌梡場巕偱偁傞Inducer of definitive endoderm 1 (IDE1 and IDE2) 傪昗揑偲偡傞5偮偺miRNA僼傽儈儕乕 (miR159, miR169, miR172, miR173, miR394) 偑丄Kong and Yang (2010) 偵傛偭偰摨掕偝傟偨丅
Yang (2010)偵傛傝丄僔儘僀僰僫僘僫偵偍偄偰摨掕偝傟偨丅
僔儘僀僰僫僘僫偱偼丄揝婹夓壓偱miR408偑僟僂儞儗僊儏儗乕僩偝傟丄偦偺昗揑乮僼僃僲儘僉僔僟乕僛偲僼僃儘僉僔僟乕僛乯偺敪尰偑堎側傝丄揝寚朢偵懳偡傞怉暔懴惈偵娭梌偡傞偙偲偑娤嶡偝傟偰偄傞乮Carrió-Seguí et al.乯
Oryza sativa偱偼丄miR11丄miR26丄miR30丄miR31偑崅揝擹搙壓偱僟僂儞儗僊儏儗乕僩偝傟傞偙偲偑娤嶡偝傟丄偦傟偼帺慠掞峈娭楢儅僋儘僼傽乕僕抈敀幙4乮NRAMP4乯傪昗揑偲偡傞偲尒傜傟偨乮Paul et al.2016). Wang傜乮2021乯偼丄僀僱偺揝寚朢壓偱miR159偲miR408偑慾奞偝傟傞偙偲傪娤嶡偟偨丅
僀僱偺揝寚朢壓偱偼丄偦傟傜偺昗揑乮ZFP丄僇儖儌僕儏儕儞條僞儞僷僋幙27丄OsGAMYB丄偍傛傃OsGAMYB乯偑夁忚偵敪尰偝傟傞寢壥偲側偭偨偙偲傪妋擣偟偨丅
摨條偵丄miR172丄miR395丄miR398丄偍傛傃miR408偼丄僗僩儗僗懴惈嫮壔丄棸墿峆忢惈偺挷愡丄摵峆忢惈偺挷愡丄摵/垷墧僗乕僷乕僆僉僔僪僨傿僗儉僞乕僛妶惈偺妶惈壔丄偍傛傃儕僌僯儞拁愊偺挷愡傪傕偨傜偡傛偆偵僟僂儞儗僊儏儗乕僩偝傟偰偄傑偟偨乮Jin et al.丄2021乯丅
丂丂
丂丂
埲壓丄恾偺愢柧丂
丂
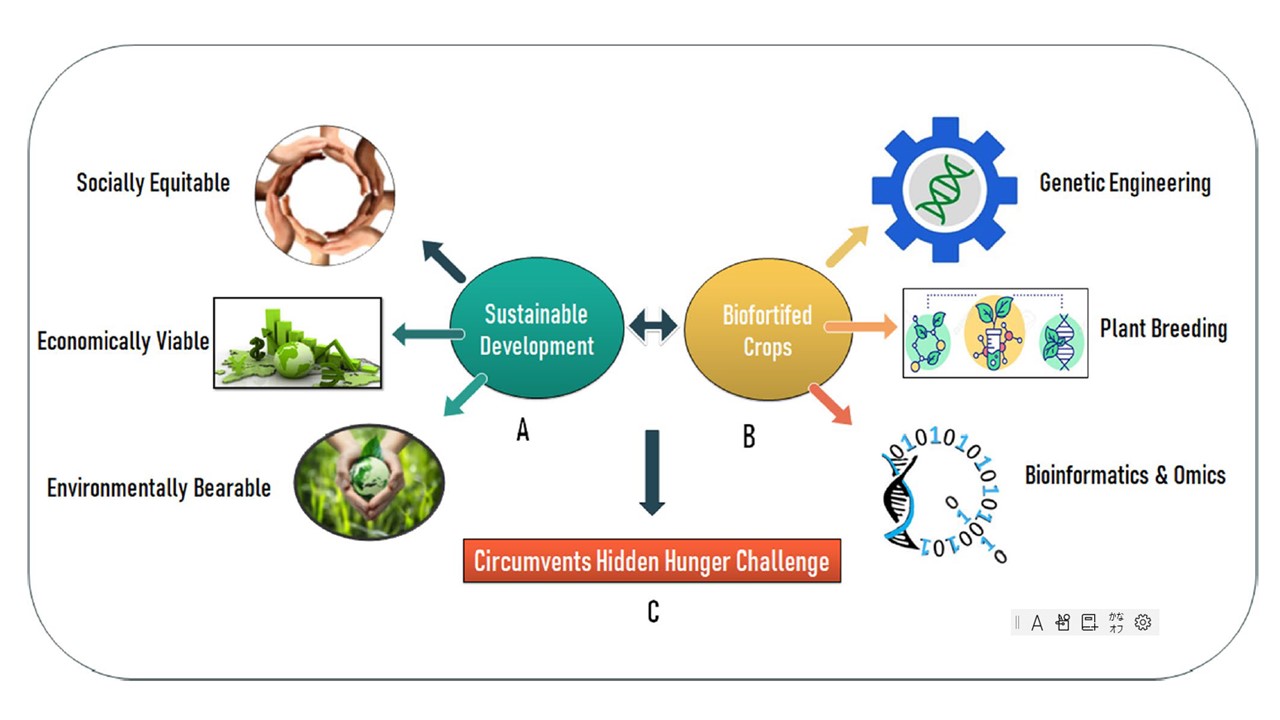
恾 1 僋儔僗僞乕恾
(A) 帩懕壜擻側奐敪偺僗僉乕儉偲丄奐敪偵偍偗傞偦偺娭學丅
(B)條乆側傾僾儘乕僠偵傛傞僶僀僆嫮壔嶌暔偺奐敪丅
miRNA傪棙梡偟偨塰梴嫮壔嶌暔偺奐敪偼丄塀傟偨婹夓栤戣傪夞旔偡傞偨傔偺怴偟偄僷儔僟僀儉偱偁傞丅
丂丂
丂丂
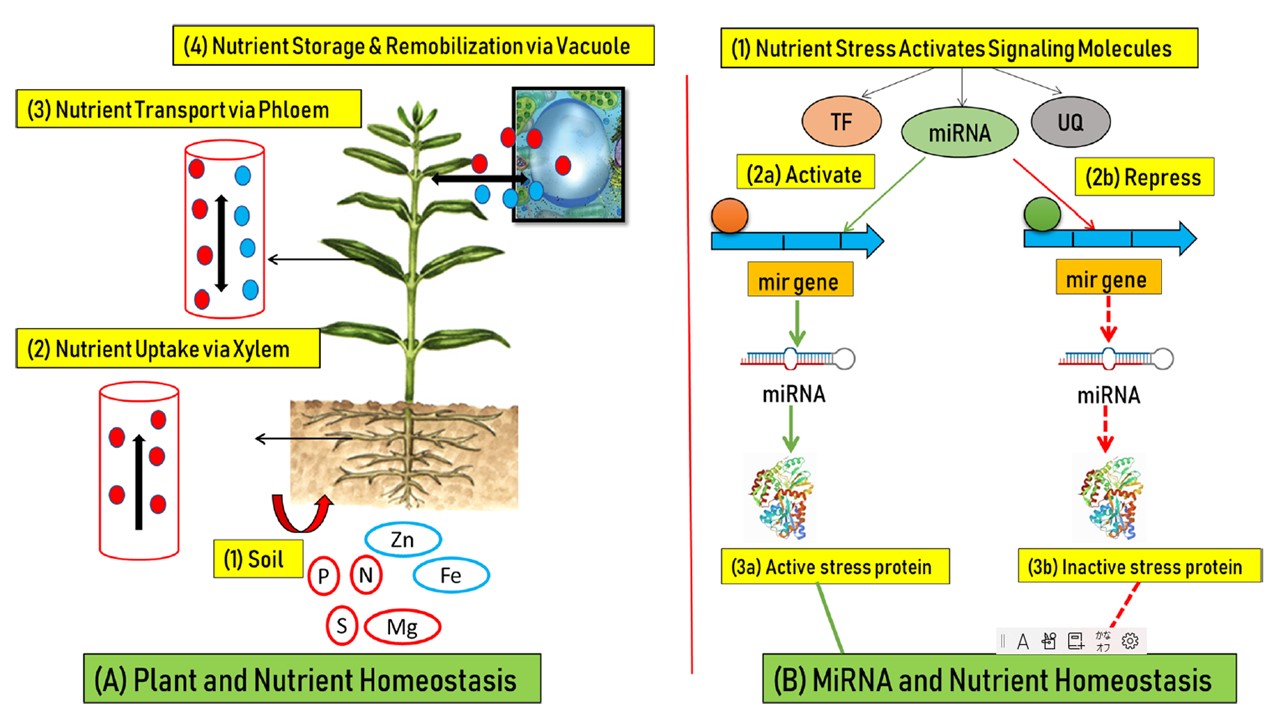
恾2 怉暔-塰梴慺-MiRNA憡屳嶌梡偺僱僢僩儚乕僋恾丅
A 怉暔偲塰梴慺偺 怉暔偲塰梴慺偺儂儊僆僗僞僔僗丗(1) 搚忞偼怉暔偑棙梡偱偒傞塰梴慺偺庡側嫙媼尮偱偁傞丅 (2)堦曽岦偵棳傟傞梴暘乮愒乯偼栘晹傪宱桼偟偰庢傝崬傑傟傞丅堐娗懇傪宱桼偟偰庢傝崬傑傟傞丅(3) 梩宻庽塼偵傛傞塰梴慺偺憃曽岦桝憲乮愒/惵乯丅(4) 塰梴慺偼偝傜偵怉暔偺塼朎傪夘偟偰挋憼乮愒乯丄嵞屌掕壔乮惵乯偝傟傞丅
B miRNA偲塰梴偺峆忢惈丗(1)塰梴晄懌偵側傞偲丄怉暔偺僔僌僫儖揱払僇僗働乕僪偼妶惈壔偟丄揮幨場巕乮TF乯丄儅僀僋儘RNA乮miRNA乯丄儐價僉僠儞傪夘偟偨僞儞僷僋幙乮UQ乯側偳偺僔僌僫儖揱払暘巕傪棙梡偟偰忣曬傪揱払偡傞丅
(2a) 僗僩儗僗僔僌僫儖偼丄miRNA偺昗揑堚揱巕傪妶惈壔偡傞偐丄(2b) 梷惂偡傞偙偲偑偱偒傞
miRNA偺敪尰偺嵎偼丄埲壓偺偄偢傟偐偺妶惈壔傪傕偨傜偡丅
(3a) 僗僩儗僗墳摎惈僞儞僷僋幙偺妶惈壔丄傑偨偼(3b)僗僩儗僗墳摎惈僞儞僷僋幙偺晄妶惈壔傪傕偨傜偡丅
偙偺傛偆偵丄miRNA偼怉暔偺塰梴僗僩儗僗帪偵億僕僥傿僽側惂屼場巕偲偟偰傕僱僈僥傿僽側惂屼場巕偲偟偰傕摥偔偙偲偑偱偒傞丅
丂
丂
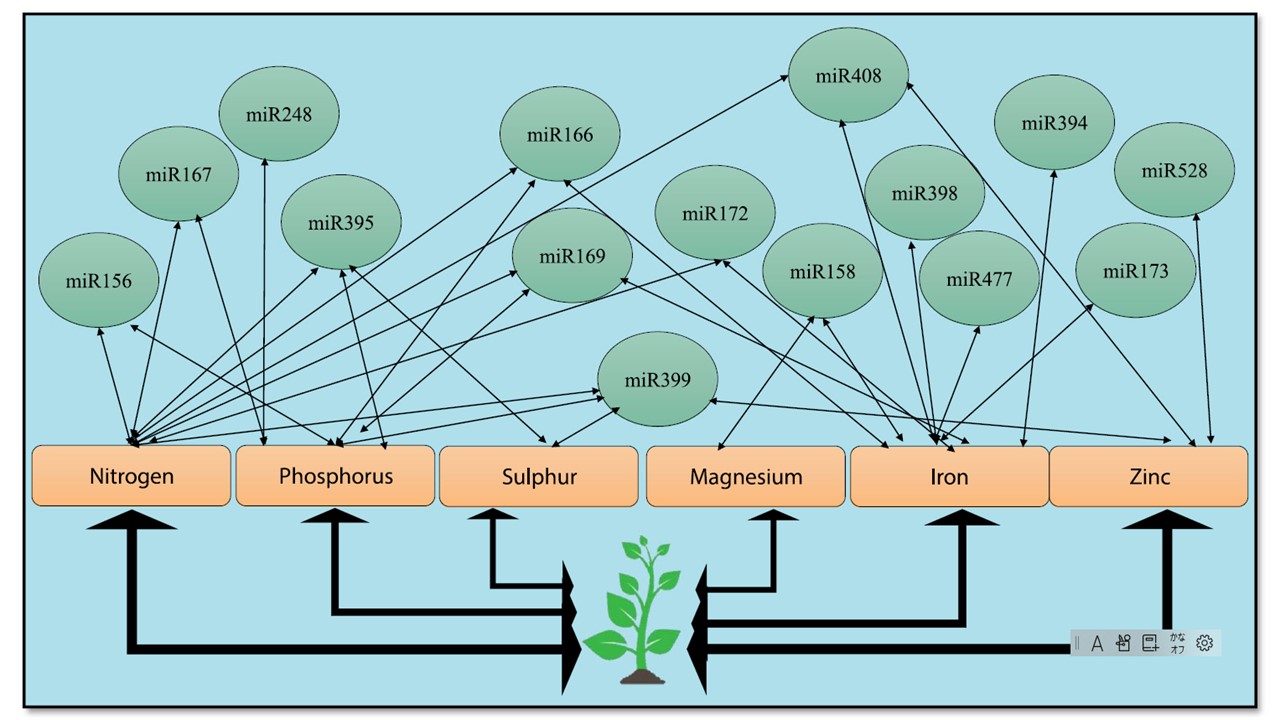
恾3 塰梴峆忢惈堐帩偵偍偗傞miRNA偺惂屼丅
埲忋丄塰梴僗僩儗僗偵墳摎偡傞miRNA傪帵偟偨丅扨堦偺miRNA偑暋悢偺塰梴慺偺峆忢惈堐帩傗媧廂偵娭梌偟偰偄傞偙偲偑曬崘偝傟偰偄傞丅
miRNA偺暋悢偺昗揑傪摨掕偟丄僶僀僆僼僅乕僥傿僼傽僀僪嶌暔偺嶌弌偵棙梡偡傞偙偲偑偱偒傞丅
Kong and Yang (2010), Liang et al. (2012), Li et al. (2013), Paul et al. (2015; 2016), Li et al. (2017c), Liang et al. (2017), Dos santos et al. (2019), Bao傜乮2019乯丄Zeng傜乮2019乯丄Liu傜乮2020a丄b乯丄Shi傜乮2020乯丄Li傜乮2020乯丄Wang傜乮2021乯丄Ding傜乮2021乯偱偁傞丅
恾1
恾2
恾3