儈僯憤愢丗儐價僉僠儞僾儘僥傾僜乕儉僔僗僥儉偲塰梴僗僩儗僗墳摎
埲壓偺僾儘僥傾僜乕儉偺婡擻偵娭偡傞儈僯儗價儏乕偱偼丄揝塰梴偵娭偡傞晹暘偺傒東栿偟偨丅
丂
丂丂丂
丂丂丂
儈僯憤愢丗儐價僉僠儞僾儘僥傾僜乕儉僔僗僥儉偲塰梴僗僩儗僗墳摎
The Ubiquitin Proteasome System and Nutrient Stress Response
Erin Mackinnon and Sophia L. Stone *
Department of Biology, Dalhousie University, Halifax, NS, Canada
Frontiers in Plant Science | www.frontiersin.org 1 May 2022 | Volume 13 | Article 867419
梫巪
怉暔偼丄僾儘僥僆乕儉偺曄壔傪懀恑偡傞儐價僉僠儞丒僾儘僥傾僜乕儉丒僔僗僥儉乮UPS乯傪娷傓條乆側暘巕婡峔傪棙梡偟偰丄惗挿傗敪払偵懳偡傞惗暔僗僩儗僗偺塭嬁傪娚榓偟偰偄傞丅
UPS偼丄摿掕偺婎幙傪儐價僉僠儞壔偟丄偦偺屻丄廋忺偝傟偨僞儞僷僋幙傪僾儘僥傾僜乕儉偱暘夝偡傞婡峔傪曪娷偟偰偄傞丅
儐價僉僠儞儕僈乕僛乮E3乯偼丄婎幙僞儞僷僋幙偵1偮傑偨偼暋悢偺儐價僉僠儞暘巕傪寢崌偝偣丄摿堎惈傪惂屼偡傞偨傔丄UPS偺拞怱揑側栶妱傪扴偭偰偄傞丅
嵟嬤偺尋媶偐傜丄UPS偼昁恵戝塰梴慺傗旝検塰梴慺偺庢傝崬傒傗桝憲偺廳梫側惂屼場巕偱偁傞偙偲偑暘偐偭偰偒偰偄傞丅
杮憤愢偱偼丄塰梴偺庢傝崬傒偲丄拏慺丄儕儞丄揝丄摵偺惂尷傑偨偼夁忚儗儀儖傪娷傓僗僩儗僗忦審偵懳偡傞墳摎偺惂屼偵娭梌偡傞E3偵偮偄偰榑偠傞丅
丂
丂
揝丗
揝偼丄僋儘儘僼傿儖偺惗崌惉丄拏慺屌掕丄DNA 偺暋惢偲廋暅丄揹巕桝憲嵔偵昁梫偱偁傞 (Pushnik et al., 2008; Zhang, 2014)丅夁忚側揝偼崻偺惉挿傪慾奞偟丄懠偺塰梴慺偺媧廂偵塭嬁傪梌偊傞偑丄偙偺栤戣偼寵婥惈丒巁惈搚忞忦審偵傛偭偰憹戝偡傞乮Becker and Asch, 2005乯丅揝偑晄懌偡傞偲丄崻偱偼揝桝憲懱1乮IRT1乯偺敪尰偑忋徃偟丄崻寳偐傜偺揝偺庢傝崬傒傪懀恑偡傞乮Vert et al.丄2002乯丅IRT1偼丄H+-ATPase2乮AHA2乯丄Fe3+傪Fe2+偵娨尦偟偰庢傝崬傓Ferric Chelate Reductase 2乮FRO2乯偲嵶朎枌偵嬊嵼偡傞暋崌懱傪宍惉偟偰丄崻寳傪巁惈壔偟偰揝暘傪壜梟壔偡傞乮Connolly et al.乯 IRT1偼丄Zn丄Mn丄僇僪儈僂儉乮Cd乯丄僐僶儖僩乮Co乯側偳懠偺昁恵梲僀僆儞傕桝憲偟偰偄傞乮Vert et al. 2002乯揝桝憲懱偼丄RING宆E3 IRT1 Degradation Factor 1乮IDF1乯偵傛偭偰嵶朎枌偱儐價僉僠儞壔偝傟丄僾儘僥傾僜乕儉偵傛傞僞乕儞僆乕僶乕偲摨條偵塼朎暘夝偵帄傞僄儞僪僒僀僩乕僔僗傪懀恑偡傞偲帵嵈偝傟偰偄傞乮Barberon et al.丄2011丟 Shin et al.丄2013丟Dubeaux et al.丄2018乯丅Zn丄Mn丄Co側偳偺旕揝嬥懏偺擹搙傪忋偘傞偲丄IRT1偺儌僲儐價僉僠儞壔偵懕偄偰丄63-儕僕儞寢崌傪梡偄偨IDF1-拠夘億儕儐價僉僠儞壔偵傛傝嵔傪惗惉偡傞偙偲偑懀恑偝傟傞乮Barberon傜丄2011; Dubeaux傜丄2018乯丅儌僲儐價僉僠儞壔偍傛傃億儕儐價僉僠儞壔偺椉曽偺廋忺偼丄塼朎偱偺撪嵼壔偍傛傃暘夝傪懀恑偡傞偙偲偵傛偭偰丄嵶朎枌偵偍偗傞IRTI偺儗儀儖傪掅壓偝偣傞丅夁忚側旕揝嬥懏偵墳摎偟偰丄IDF1傪夘偟偨儐價僉僠儞壔傊偺愗傝懼偊偼丄IRT1偺CIPK23埶懚揑側儕儞巁壔偵傛偭偰堷偒婲偙偝傟傞乮Dubeaux et al.丄2018乯丅AHA2偍傛傃FRO2傕儐價僉僠儞壔偝傟傞偑丄偙偺廋忺偼旕揝嬥懏偵傛偭偰桿摫偝傟偢丄撪嵼壔偍傛傃暘夝傪懀恑偟側偄偑丄峺慺婡擻傪挷愡偡傞偙偲偑帵嵈偝傟偰偄傞乮Martín-Barranco et al, 2020乯丅FER條揝寚朢桿敪揮幨場巕乮FIT乯偼丄IRTI傗懠偺揝寚朢墳摎惈堚揱巕偺敪尰傪惂屼偡傞婎杮僿儕僢僋僗儖乕僾僿儕僢僋僗乮bHLH乯揮幨場巕孮偺拞偺廳梫側惂屼場巕偱偁傞乮Cohen傜丄2004丟Mai傜丄2016乯丅揝寚朢偼丄FIT偺敪尰偲妶惈壔傪捀揰偲偡傞bHLH惂屼場巕偺僇僗働乕僪傪堷偒婲偙偟丄揝偺摦堳偍傛傃庢傝崬傒偵娭梌偡傞堚揱巕偺揮幨傪懀恑偡傞丅2偮偺bHLH揮幨場巕丄bHLH105偲bHLH115偼丄RING宆E3 BRUTUS乮BTS乯偵傛偭偰儐價僉僠儞壔偝傟僾儘僥傾僜乕儉暘夝偺昗揑偲側傝丄僗僩儗僗偺側偄忬懺偱揝寚朢斀墳偺妶惈壔傪嬛巭偡傞乮Selote et al.丄2015擭乯丅Iron Mans乮IMA乯偼丄Fe寚朢壓偱BTS偵寢崌偟丄bHLH105偍傛傃bHLH115偲偺憡屳嶌梡傪攋夡偟丄揮幨場巕傪拁愊偝偣偰僗僩儗僗墳摎傪懀恑偡傞儁僾僠僪僼傽儈儕乕偱偁傞乮Grillet et al., 2018; Li et al., 2021乯丅BTS偼丄BRUTUS-LIKE1乮BTSL1乯偍傛傃BTSL2傪娷傓僿儊儕僩儕儞娷桳RING宆E3偺僼傽儈儕乕偵懏偟丄O. sativa Hemerythrin motif-containing RING- and Zinc-finger protein 1乮OsHRZ1乯偍傛傃 OsHRZ2乮彫椦傜丄2013; Rodríguez-Celma et al, 2019乯偵椶帡偟偰偄傞丅BTSL1偍傛傃BTSL2偼丄僾儘僥傾僜乕儉埶懚惈暘夝傪夘偟偰FIT偺懚嵼検傪挷愡偡傞乮Rodríguez-Celma et al.丄2019乯丅BTS丄OsHRZ1丄偍傛傃OsHRZ2偼丄Fe偍傛傃Zn偲寢崌偡傞偙偲偑帵偝傟偰偍傝丄E3偑嬥懏僙儞僒乕偲偟偰偩偗偱側偔丄Fe寚朢墳摎偺晧偺儗僊儏儗乕僞乕偲偟偰婡擻偡傞偙偲偑帵嵈偝傟偰偄傞乮Kobayashi et al.乯 OsHRZ1偍傛傃OsHRZ2偼傑偨丄夁忚側揝僗僩儗僗壓偱嬥懏偺庢傝崬傒傪惂尷偡傞偨傔偵昁梫偱偁傝丄揝姶抦偵偍偗傞E3s偺栶妱傪偝傜偵帵嵈偟偰偄傞乮Aung傜丄2018擭乯丅RING宆E3偱偁傞Ring Domain Ligase1乮RGLG1乯偍傛傃RGLG2傕丄揝惂尷忦審傊偺墳摎偺惂屼偵娭梌偟偰偄傞偑丄婎幙偼晄柧偱偁傞乮Pan et al.丄2015乯丅
丂丂
丂丂
寢榑
揝傗拏慺側偳偺塰梴慺偺峆忢惈堐帩偵娭傢傞儐價僉僠儞傪夘偟偨僾儘僙僗偵娭偡傞抦幆偼廫暘偵妋棫偝傟偰偄傞丅UPS偼丄崻寳偐傜塰梴傪庢傝崬傓嵶朎枌偺僩儔儞僗億乕僞乕偺懚嵼傗丄崻偐傜抧忋婍姱偵塰梴傪堏峴偝偣傞僩儔儞僗億乕僞乕偺検傪挷愡偟偰偄傞丅傑偨丄UPS偼丄塰梴偺峆忢惈堐帩偵娭傢傞堚揱巕偺揮幨傪惂屼偡傞惂屼場巕偺儗儀儖傕挷愡偟偰偄傞丅偟偐偟丄儂僂慺丄垷墧丄摵側偳丄傎偲傫偳偺塰梴慺偺庢傝崬傒傪惂屼偡傞UPS偺婡擻偵偮偄偰偺棟夝偼丄旕忢偵尷傜傟偰偄傞丅尋媶偑恑傔偽丄儐價僉僠儞傪夘偟偨塰梴慺偺姶抦丄庢傝崬傒丄堏摦偺惂屼偺斖埻偑傛傝柧傜偐偵側傞偙偲偼娫堘偄側偄丅傑偨丄儐價僉僠儞儕僈乕僛偺妶惈傗婎幙偲偺寢崌傪丄偝傑偞傑側塰梴忦審壓偱惂屼偡傞儊僇僯僘儉偵偮偄偰傕丄傛傝怺偄棟夝偑摼傜傟傞偐傕偟傟側偄丅婥岓曄摦傪峫椂偡傞偲丄塰梴慺偺妉摼偵偍偗傞儐價僉僠儞壔偺栶妱傪傛傝徻嵶偵抦傞偙偲偼丄惗堢傗廂妌偵懳偡傞惗暔揑丒惗暔妛揑僗僩儗僗偺埆塭嬁傪夵慞偡傞偨傔偵丄嵟揔側塰梴忬懺傪堐帩偡傞曽朄偺棟夝偵栶棫偮偲巚傢傟傞丅
丂丂
丂丂 丂
昞1丂塰梴慺媧廂偲僗僩儗僗墳摎偵娭傢傞儐價僉僠儞峺慺孮
丂
恾1乥乮A乯儐價僉僠儞丒僾儘僥傾僜乕儉宯偺娙棯壔偟偨奣梫丅E1丄E2偍傛傃E3峺慺偼丄1偮傑偨偼暋悢偺儐價僉僠儞乮Ub乯暘巕偺昗揑婎幙傊偺寢崌傪懀恑偡傞丅昗揑偺儐價僉僠儞壔偼丄婎幙偵寢崌偡傞慜偵E2偐傜E3妶惈僔僗僥僀儞偵Ub偑堏摦偡傞乮i乯偐丄婎幙偵Ub偑捈愙堏摦偡傞乮ii乯偙偲偵傛傝峴傢傟傞丅偙偺寢崌僇僗働乕僪偵傛傝丄婎幙偺儌僲儐價僉僠儞壔乮1僇強偵1偮偺Ub乯丄儅儖僠儐價僉僠儞壔乮堎側傞晹埵偵暋悢偺Ub乯丄億儕儐價僉僠儞壔乮暋悢偺Ub偑嵔忬偵側傞乯偑惗偠傞丅億儕儐價僉僠儞壔偝傟偨婎幙偼丄26S僾儘僥傾僜乕儉偵傛偭偰擣幆偝傟暘夝偝傟傞丅Ub偼婎幙偐傜彍嫀偝傟丄嵞棙梡偝傟傞丅(B)堎側傞E3僞僀僾偺柾幃恾丅E3偼RING (Really Interesting New gene)丄HECT (Homologous to E6AP C-terminus)丄U-box僪儊僀儞傪棙梡偟偰E2偲憡屳嶌梡偟偰偄傞丅僔儞僌儖僒僽儐僯僢僩偺HECT傗RING-in-between-RING乮RBR乯僞僀僾偺E3偼丄E2偐傜Ub傪庴偗庢傞丅暋崌宆E3 Culin (Cul)-RING 儐價僉僠儞儕僈乕僛 (CRLs) 偼丄E2偲婎幙偲偺憡屳嶌梡偵堎側傞僒僽儐僯僢僩傪棙梡偡傞丅
丂
丂
恾2乥僔儘僀僰僫僘僫偲僀僱桼棃偺E3偑塰梴愛庢傪惂屼偡傞栶妱傪娙棯壔偟偨恾丅(A)崅拏慺壓偱偼丄E3 NLA偑儐價僉僠儞埶懚揑偵NRT1.7徤巁僩儔儞僗億乕僞乕傪暘夝偟丄拏慺偺夁忚拁愊傪夞旔偟偰偄傞丅僩儔儞僗儗僙僾僞乕NRT1.1B偼E3 NBIP1傪摦堳偟丄SPX4傪儐價僉僠儞壔偟偰揮幨場巕NLP3偑妀撪偵擖傝丄拏慺墳摎惈堚揱巕偺敪尰傪懀恑偝偣傞偙偲偑傢偐偭偨丅(B乯儕儞巁偑朙晉側娐嫬偱偼丄E3 NLA偑柍婡儕儞巁桝憲懱PHT1傪儐價僉僠儞壔偟丄26S僾儘僥傾僜乕儉偵傛傞暘夝傪懀恑偟丄儕儞巁偺庢傝崬傒傪梷惂偟偰夁忚拁愊傪杊偄偱偄傞丅儕儞巁惂尷僗僩儗僗忦審壓偱偼丄E3偱偁傞SDEL1偲SDEL2偑SPX4偺暘夝傪拠夘偟丄揮幨場巕PHR1/2偑PHT1側偳偺PSI堚揱巕偺敪尰傪妶惈壔偡傞偙偲偑偱偒傞丅傑偨丄E3偱偁傞PRU1偑梷惂場巕WRKY6偺暘夝傪拠夘偟丄PHO1偺揮幨偺梷惂傪娚榓偟偰偄傞丅PHO1僩儔儞僗億乕僞乕偺懚嵼検偑憹壛偡傞偲丄崻偺栘晹傊偺Pi偺儘乕僪偑懀恑偝傟傞丅
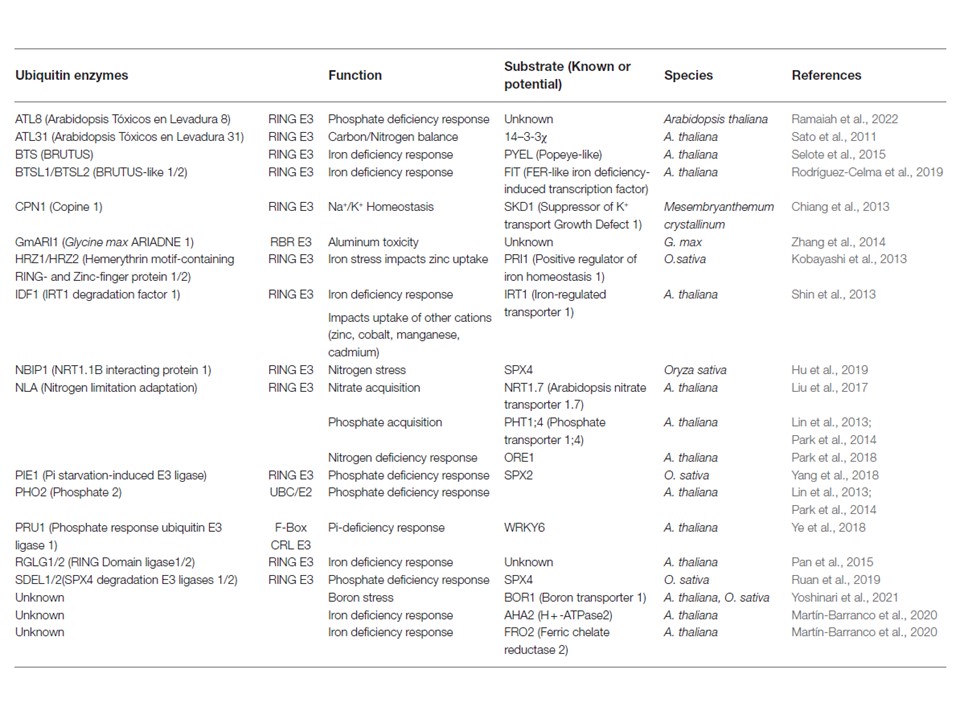
昞1
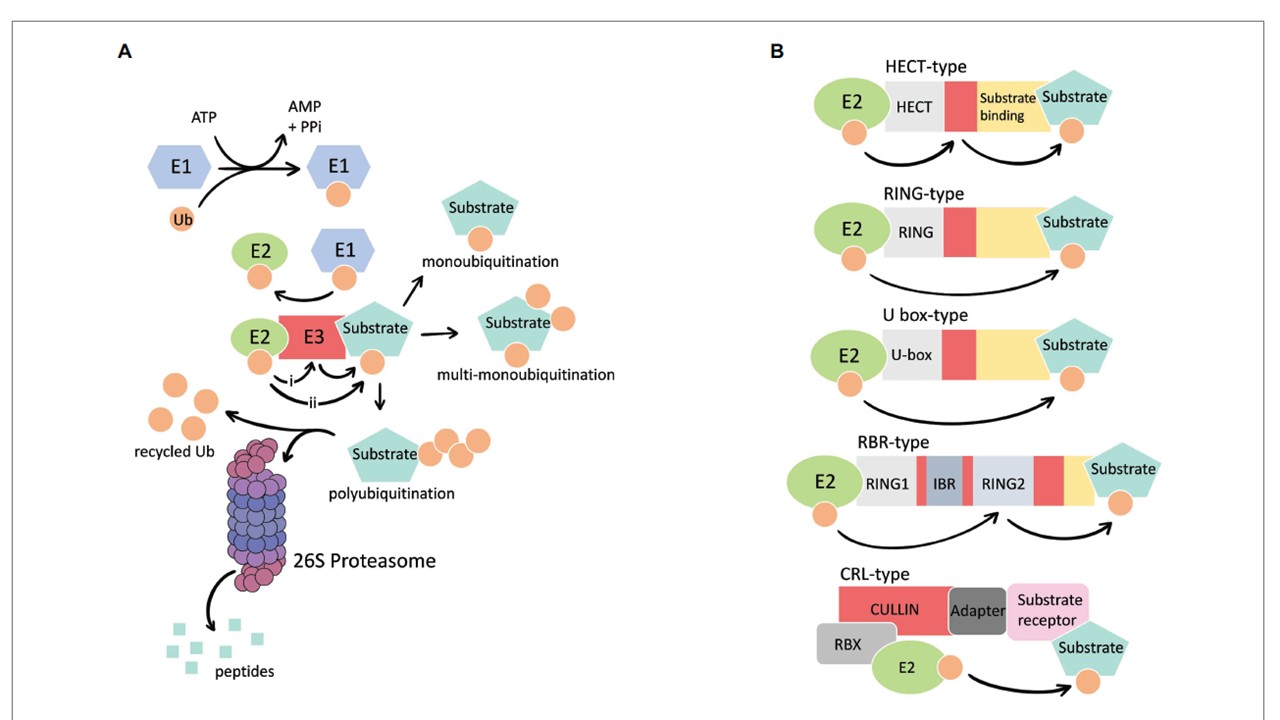
恾1
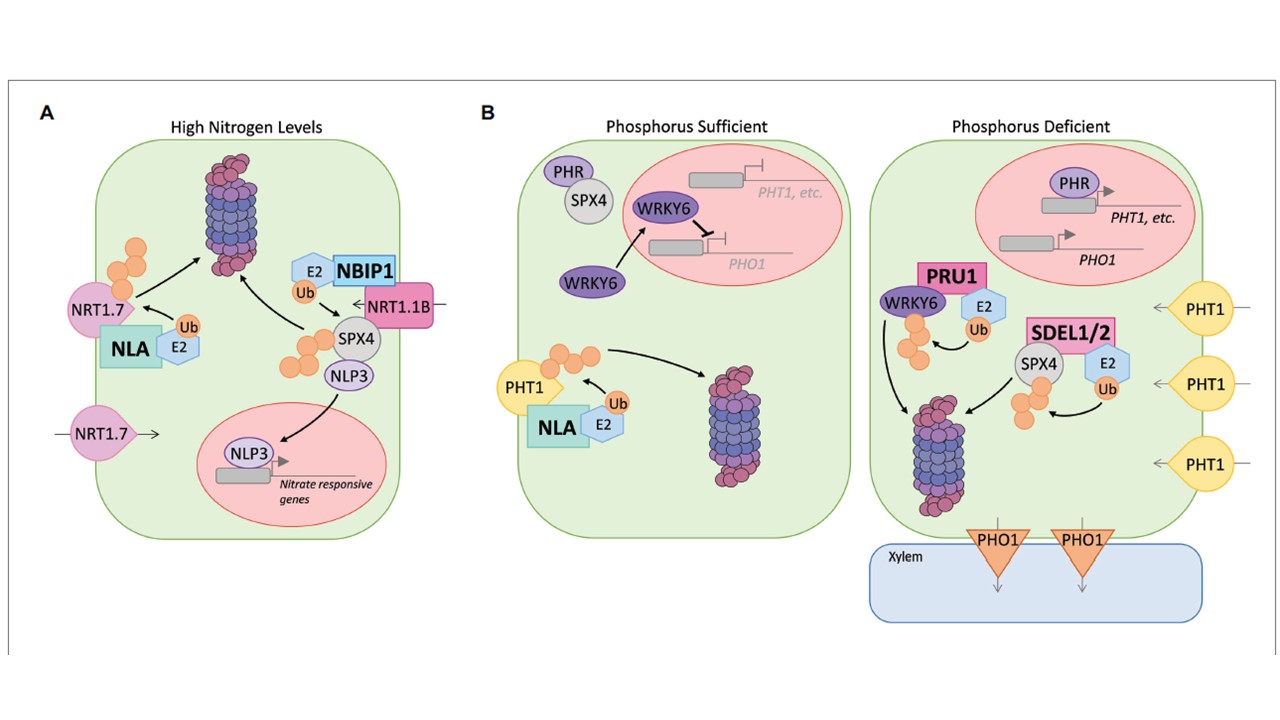
恾2