干ばつはミネラル栄養を特異的にダウンレギュレートする。植物イオンの含有量と関連する遺伝子の発現
この論文は、干ばつ時に、各種の元素イオンやその元素に関連する遺伝子発現がどのように変動するかを総合的に調べたものである。
図7のうち、小生が興味がある鉄イオンに関連する遺伝子発現の部分を図7’として拡大して、提示しておいた。
干ばつはミネラル栄養を特異的にダウンレギュレートする。植物イオンの含有量と関連する遺伝子の発現
Aurélien D'Oria、Galatéa Courbet、Bastien Billiot、Lun Jing、Sylvain Pluchon、Mustapha Arkoun、Anne Maillard、Christine Paysant-Le Roux ...
初出:2022年8月5日
https://doi.org/10.1002/pld3.402
Plant Direct Volume 6, Issue 8 e402
(要旨)
植物収量の主な制限要因の一つは干ばつであり、この環境ストレスに対する生理的反応は広く報告されているが、ミネラル栄養への影響に関する研究はほとんどない。Brassica napusとTriticum aestivumを中度あるいは重度の水不足に曝し、機能的イオノミック解析と20種類の栄養素の純吸収量の算出によって、干ばつに対する応答を評価した。ほとんどの養分摂取量は減少したが,Fe,Zn,Mn,Mo の摂取量は,評価した多くの生理的パラメータ (成長,ABA 濃度,ガス交換,光合成活性)よりも早く,かつ大きなスケールで影響を受けていることが わかった。さらに、B. napusにおいて、イオノームに関連する遺伝子(Known ionomic genes, KIGs)、あるいは特定の栄養分の輸送に関与すると想定される183の葉における差次的発現遺伝子のパターンを解析した。その結果、干ばつ時の遺伝子発現は、アップ(ClとCoの輸送)、ダウン(N、P、B、Mo、Niの輸送)、あるいは混合レベル(S、Mg、K、Zn、Fe、Cu、Mnの輸送)の調節からなる3つのパターンがあることが明らかになった。この3つの遺伝子制御パターンについて、特定の遺伝子機能、葉のイオン組成の変化と関連づけ、要素間で確立されたクロストークを考慮しながら議論した。観察されたFeの取り込み減少は、乾燥に対する特異的な反応を介して起こり、間接的にZnとMnの取り込み減少につながり、これらはダウンレギュレーションされた遺伝子にコードされる共通のトランスポーターによって取り込まれる可能性が示唆された。
(はじめに)(常識的な部分を排して部分訳した)
ここ数十年、「-omic」ハイスループット技術が採用され、遺伝子発現(「トランスクリプトミクス」)、代謝物(「メタボロミクス」)または元素組成(「イオノミクス」)を同時に特定および定量し、生物または生物ストレスに応答した代謝変化を解読することができるようになった。これらのアプローチにより、異なる生物における1つの元素または元素群の蓄積を制御する遺伝子の同定に成功している。最近、Whittら(2020)は、植物のイオノームに影響を与える遺伝子のキュレーションリストを提案し、「Known Ionomic Genes」(KIGs)と称した。この遺伝子リストは、当初、KOまたは過剰発現変異体がイオノームの少なくとも1つの要素について影響を受けたことを示す公開著作物の利用可能性に応じて、シロイヌナズナ(これまでに同定された136遺伝子)から確立され、その後、他の種に拡張された(10種は1588のオルソログを明らかにしている)。含まれる遺伝子は、主にイオン輸送とホメオスタシスに関与し、二次的に金属イオンキレートや他のタイプの輸送やイオンへの反応に関与している可能性がある。しかしながら、このKIGリストは利用可能な知識によって制限されており、著者らによれば、トランスポーター遺伝子や鉄や亜鉛の蓄積の変化に関与する遺伝子が過剰に含まれているとのことである。対照的に、MgとCaに関与する可能性があり、主にNの輸送に関連する遺伝子は、このKIGリストにはあまり含まれていない。このリストは、生物学的ストレスがイオンの組成にどのように影響するかを解読するのに有効であるが、イオン輸送に関連する遺伝子を遺伝子オントロジーで濃縮することで、より完全な情報を提供できるようになると思われる。
そこで本研究では、水分制限の因果関係を明らかにするために、農学的に注目されている対照的な2つの植物種(B. napus および T. aestivum)において、(i)成長、および水ストレスの生理的・形態的指標(画像解析、ガス交換、13C 識別、植物ホルモン蓄積)と(ii)根への純吸収および機能性イオノームを形成する 20 要素の組織内容に対する中程度または重度の乾燥の影響を評価することを目的とした。さらに、B. napusにおいて、Whittら(2020)が以前に確立したKIGリストの遺伝子の発現パターンに照らし合わせ、Gene Ontologyから得られた「イオン輸送」の注釈付き遺伝子で拡張して、乾燥によって誘発される葉イオノームの変化を検討した。そして、関連する遺伝子の関与を、干ばつによって引き起こされる栄養摂取量とイオン組成の変化に照らして解釈し、これまでに報告されたミネラル栄養不足の影響に従って議論した。
(論議)(最後のパートの一部のみ訳した)
4.3 栄養素吸収量の減少とそれに伴うイオノミック組成の変化は、輸送に関わる遺伝子発現の変化で説明できる。
干ばつ期間または強度が増加すると、ほとんどの元素含有量が大きく減少し、遺伝子発現が変化することが明らかになった。イオノミック組成に基づいて、特定の栄養欠乏下での輸送遺伝子制御に関する現在の知識でこれを解釈することは魅力的なことである。
一般に、栄養トランスポーター、同化酵素、転写または調節因子をコードする遺伝子の発現は、通常、栄養欠乏に伴って上昇し、逆に栄養充足下では下降することが知られている。例えば、N (Krapp et al., 2014), S (Sultr1.1; 1.2; Courbet et al., 2019; Shinmachi et al., 2010), K (Ashley et al., 2006; Wang et al., 2013), またはFe (Irt1, Vert et al., 2002)がそうである。
我々の干ばつ条件下では、Sの取り込み、およびS組織含量が強く減少し、 Sのホメオスタシスを制御する核局在タンパク質MSA1の発現増加(Huang et al, 2016)、SO42-トランスポーターをコードする遺伝子(Sultr3.1, 3.4, 4.1)の発現増加との関連性が見られた。一方、水制限植物では葉中のN含量は安定したまま(または増加した)。おそらく旧葉から若葉へのNの再移動が増加した結果であり、若葉のNトランスポーター(AMT1.1, AMT1.2, AMT2, DUR3, NRT2.5; NRT3.1, NFP6.3, NFP7.3)のコードをもつ遺伝子発現の誘導は必要ないものと考えられる。鉄の場合は、干ばつによってその取り込みが強く減少し、その結果、すべての組織でその 含有量が大きく減少したので、より興味をそそられる。文献(Eide et al., 1996; Henriques et al., 2002; Nishida et al., 2012)によると、このようにFe含量が強く減少すると、IRT1の発現が増加するはずであると予想された。しかし、菜種の芽を対象に行った IRT1 RT-Q PCR 解析(データなし)では、水ストレス植物と対照植物の間で IRT1 の発現に有意な差は認められなかった。このことは、鉄欠乏時に想定されるような干ばつ時の IRT1 発現による鉄獲得制御(根および葉で強 く増加)は行われていないことを示しており、干ばつ時には他の特異的な機構が関与している可能性を示 唆するものである。イオノミック側では,いくつかの研究により,Feと他の栄養素の間に多数のクロストークが存在することが明らかにされており(Billard et al., 2014; Courbet et al., 2019; Forieri et al., 2013; Maillard, Sorin, et al., 2016; Vigani & Briat, 2016),これらの相互関係の中にはFeのみならず他の金属も輸送できる取込系が含まれている。実際、これらの遺伝子のいくつかは、短期および長期の厳しいWDまたは長期の中等度のWDへの曝露後にダウンレギュレートされることがわかった。
この遺伝子群の中では、(i)Fe、Zn、Mnを輸送する可能性のあるNRAMP1(Cailliatteら、2010);(ii)Fe、Zn、Mn、Cuなどのリガンドや金属を輸送することが知られているYSL1(Watersら、2006);(iii)ZIF1(Haydonら、2012)金属リガンドトランスポーター;(iv) 鉄のホメオスタシスに関与するとされ、その KO 変異体が鉄蓄積を示す FRD3/MAN1(Rogers & Guerinot, 2002)などが知られている。
従って、干ばつ下では、遺伝子発現の低下により鉄の取り込みが抑制され、二次的に Mn、Cu、Zn の取り込みが抑制されることが示唆された。その後、すなわち長期間の厳しい干ばつの後、低い葉の Fe 濃度は、Fe に関する遺伝子の発現上昇と関連している。これは、シロイヌナズナで過剰発現させると地上部の Fe 濃度が高くなることが示されている ZIF1 や (Haydon et al., 2012) 、Fe 欠乏下でシロイヌナズナの成長を正に制御することが報告されている転写因子 PYE がそうである (Long et al., 2010) ことから明らかであった。
最後に、別の研究(Rasheed et al., 2016)は、土壌で栽培されたシロイヌナズナが水分制限後の根でIRT1、IRT3、FRO2などのFeの取り込みに関わる遺伝子が早期にダウンレギュレートすることを示し、乾燥によってFe輸送と代謝が特異的に影響を受けるという我々の観察を裏付けている。
現段階では、本研究で乾燥時に観察されたMo取り込みの特異的なダウンレギュレーションは、MOT1遺伝子の発現が低下した結果であること以外に、どのような制御が行われているかを説明するのはより困難である。
図7の説明。 その次の図7‘は図7の鉄の部分のみを拡大したもの。
輸送関連遺伝子と既知のイオノミック遺伝子がWD処理によって同時にアップレギュレートおよびダウンレギュレートされた発現パターン。差次的発現遺伝子(DEGs)のLog2 fold変化は、調整p値<.05にアスタリスク「*」を付けて表示した。各パネルの上部にあるグラフは、各遺伝子セットに関連する要素の相対的な含有量(濃いグレー、有意、薄いグレー、非有意)の変動を表している
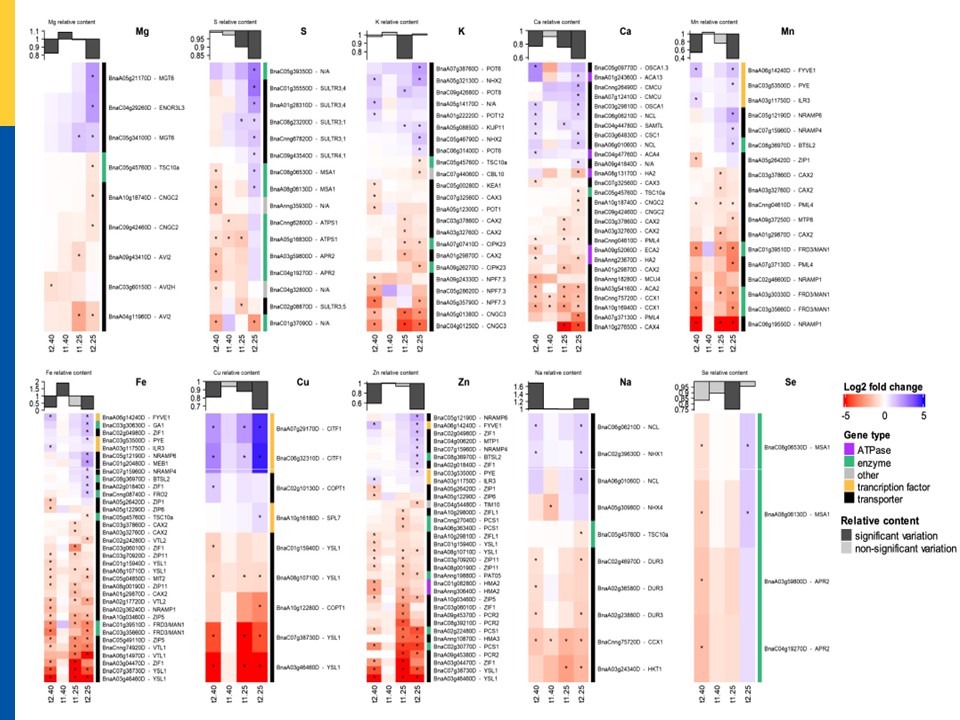
Fig7

Fig7'