(dmas関連論文)トウモロコシAldo-Keto還元酵素-4ファミリーに関する機能ゲノミクスと構造的知見:胚におけるストレス代謝と基質特異性
以下の論文に出てくるdmas(deoxymugineic acid synthase)遺伝子の発見は
Bashir, K., Inoue, H., Nagasaka, S. et al (2006) Cloning and characterization of
deoxymugineic acid synthase genes from graminaceous plants. J Biol Chem, 281,
32395–32402.
によるものです。
一方、ここで紹介する著者たちは、植物のaldo-keto反応について興味を持って研究しているのですが、その中で、トウモロコシ(Zea mays)の中に8種類のdmas類似遺伝子を見出して、それらのmRNA含量とたんぱく含量を測定しています。またそのうちの3種類の遺伝子についてトウモロコシ種子における部位別mRNA発現量を調べています。
Bashirさんたちはイネの根と葉でdmasの遺伝子発現をpromoter-GUSを用いて染色して見ています。すでに、19年前になりますが。
というわけで、この論文ではdmasに関する部分のみを紹介します。
Functional Genomics and Structural Insights into Maize Aldo-Keto Reductase-4
Family: Stress Metabolism and Substrate Specificity in Embryos
トウモロコシAldo-Keto還元酵素-4ファミリーに関する機能ゲノミクスと構造的知見:胚におけるストレス代謝と基質特異性
シルビア・モライス・デ・ソウザ;プリシラ・オリベイラ・デ・ジュゼッペ;マリオ・ティアゴ・ムラカミ;ジャン・チョウ・グアン;ジョナサン・W・サンダース;エドゥアルド・キヨタ;マルセロ・レイテ・サントス;エリック・A・シュメルツ、ホセ・アンドレス・ユネス;カレン・E・コック
責任著者:sylvia.sousa@embrapa.br; kekoch@ufl.edu
要旨
アルドケトレダクターゼ(AKR)は自然界に広く存在し、単糖類から潜在的に毒性のあるアルデヒドまで、幅広い基質を還元する能力を有する。植物において、AKRは反応性アルデヒドの解毒を含む主要な代謝過程に関与している。本研究の目的は、i) アルドケト還元酵素-4(AKR4s)をコードするトウモロコシ遺伝子ファミリーの解明、ii) それら間の配列と機能のギャップを埋めること、iii) 胚特異的ストレス代謝に関与するファミリーメンバーに焦点を当てることである。我々はゲノムワイド解析アプローチを用いてAKR4をコードするトウモロコシ遺伝子を同定し、3つのサブグループに分類される15メンバーの遺伝子ファミリーを定義・注釈した。発現プロファイリングと実験室検証により、以下の機能的役割が明らかになった:i) AKR4C Zm-1はストレス時のアルデヒド解毒に関与、ii) AKR4C Zm-2は多様な基質親和性を持つストレス応答性AKRを含む、iii) AKR4A/B Zm-3は鉄輸送用フィトシデロフォアなどの特殊代謝物生成に寄与。機能への配列変異の影響を調査するため、AKR4C Zm-1の代表であるZmAKR4C13を特性評価した。そのmRNAとタンパク質は主に胚に局在し、特殊な役割を示唆した。組換えZmAKR4C13はメチルグリオキサールや低分子アルデヒドを効率的に還元したが、4炭素より大きなアルドースに対しては活性が低かった。結晶構造解析により、活性部位におけるサイズ制約が同定された。これは294番位置にあるより大きなLEU残基に起因する。総合的に、我々の結果は活性部位構造の微妙な変化がAKRの基質特異性に及ぼす影響を強調している。また、トウモロコシのZmAKR4C13がメチルグリオキサールやその他の小分子代謝物の解毒に関与する可能性を示しており、これらは胚におけるストレスシグナル伝達に寄与し得る。
図1. 機能的に定義された他植物種のオルソログとのトウモロコシAKR4の系統解析。トウモロコシAKRは太字の赤色で示され、3つのサブファミリーに分類される。AKR4C Zm-1グループ(ピンク)はアルドース還元酵素であり、トウモロコシのZmAKR4C13を含む。AKR4C Zm-2グループ(緑)はトウモロコシのAKR4C7のようなケト還元酵素を含む。AKR4A/B Zm-3(青)にはトウモロコシDMAS(AKR4B6)および複雑な二次代謝産物生合成に関わるAKRが含まれる。配列アラインメントはClustalXバージョン1.83(Thompson et al., 1997)およびMUSCLE 3.7(Robert 2004)を用いて実施した。
系統関係は、LIRMMのPhyML v. 2(http://phylogeny.lirmm.fr)(Dereeper and Guignon et al., 2008)を用いて決定した。最大尤度法と近似尤度比検定により分岐支持値の統計的評価を行った。系統樹はiTOL v. 6.5.1
(Itol.embl.de) (Letunic and Bork, 2021)を用いて可視化した。起源種、機能解析、遺伝子識別子、
およびアクセス番号については表Iを参照のこと。
図2. トウモロコシAKR4遺伝子ファミリーのトランスクリプトームとプロテオームの比較。データはWalleyら(2016)から収集され、qTeller(qteller.maizegdb.org)を通じてアクセス可能。対照サンプルを用いてmRNA(FPKM、RNAseq)およびタンパク質(NSAF)の量を定量化。左列には、タンパク質および酵素名(AKR命名法で定義されている場合)、遺伝子座(MaizeGDB.orgに基づく)、およびトウモロコシゲノムバージョン5にリンクされた遺伝子識別子が示されている。背景色は図1および表1と同様に系統群1〜3を示す。Y軸の目盛りは各ファミリーメンバーごとに調整されており、特に存在量が著しく変動するタンパク質では顕著である。X軸には組織起源が表示され、図下部に拡大表示されている。SAM = 頂端分裂組織。
図4. トウモロコシ穀粒におけるZmAkr4C13 mRNAの発現。A. 穀粒発育過程における相対発現量。B. 12 DAP穀粒におけるAspergillus flavusによるZmAkr4C13の発現誘導:無損傷対照(Ctrl)、機械的損傷(Injur)、Aspergillus flavus(Inoc)接種。
C. 24系統のNAM系統における発育中穀粒でのZmAkr4C13発現のタイミングと発現量に関する遺伝的変異。これらの系統のうち5系統は
発育全期間を通じて一貫して高いZmAkr4C13 mRNAレベルを示し、Oh43系統は穀粒充填期のピーク時(20〜25 DAP)における発現上昇が顕著であった。
ここに認められる遺伝的変異は、育種によるZmAkr4C13発現増強の可能性が高いことを示唆する。
RNA-seqデータはDiepenbrock et al. (2017) から、Maize Genomics Resources (http://maize.uga.edu/gene_expr_analyses_download.shtml) 経由でダウンロード後、統合した。図AおよびB:エラーバーは3つの独立した生物学的反復実験における平均値の標準誤差(SEM)を示す。
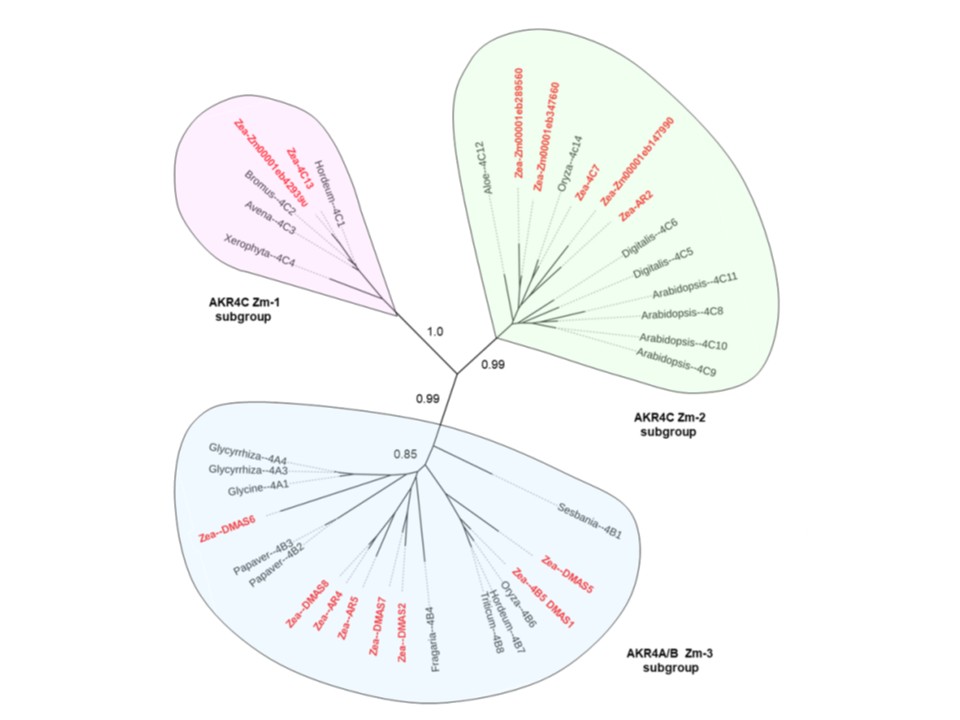
図1。機能的に定義された他植物種のオルソログとのトウモロコシAKR4の系統解析。
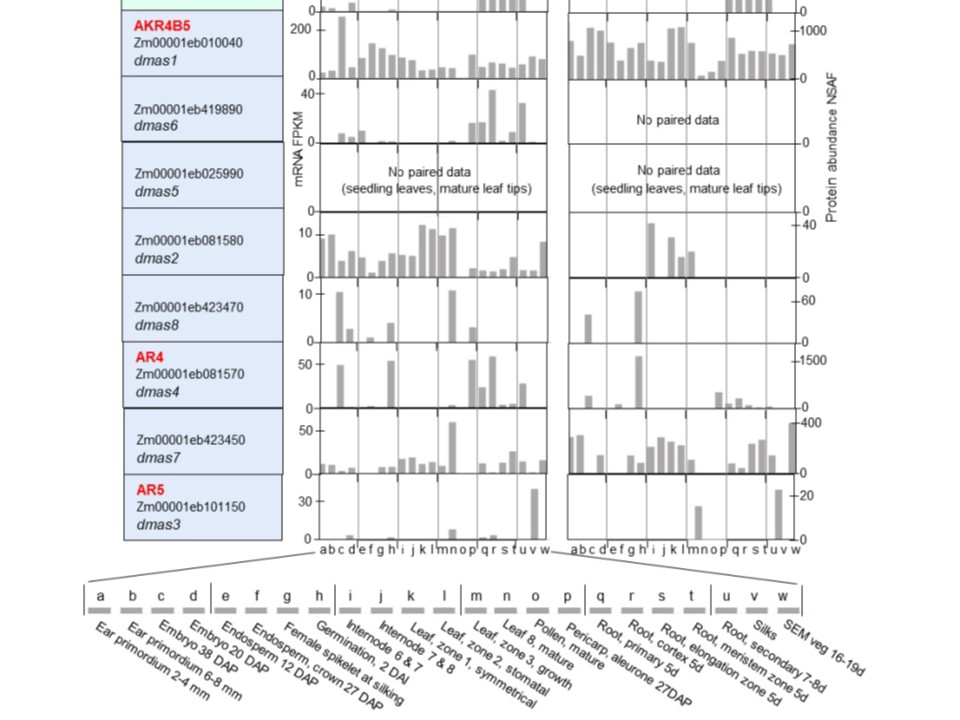
図4。トウモロコシ穀粒におけるZmAkr4C13 mRNAの発現。
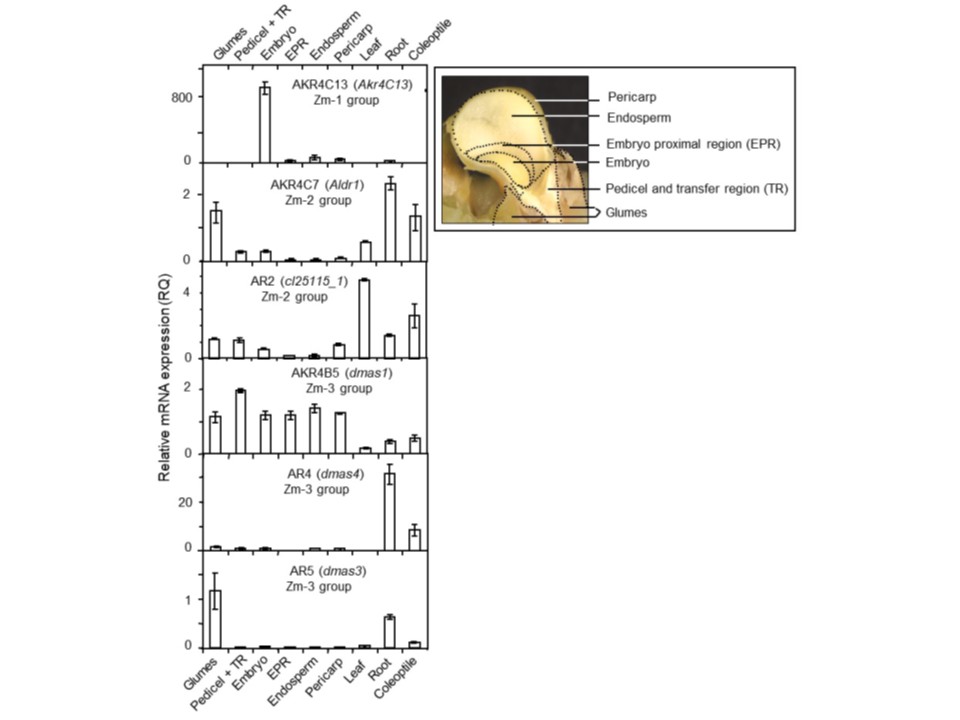
図2。トウモロコシAKR4遺伝子ファミリーのトランスクリプトームとプロテオームの比較。