穀物作物におけるbHLH転写因子:成長・発生・ストレス応答調節における多様な機能
以下の報文は、全く実験を伴わずに、文献を詳細に総合的に検索して組み立てた、総説である。小生には、鉄栄養に関する部分だけしか理解できていないが、全体的に良く書けていると思う。この論文のように既存のデータのみ(dry)を駆使して、一つの仮説を提供してみる論文を(wetなしでも)作成することも今後は評価すべきかもしれない。この論文を読んで、あらためて植物の鉄栄養に関しても、中国人研究者のレベルが格段に上がっていることを実感させられた。
穀物作物におけるbHLH転写因子:成長・発生・ストレス応答調節における多様な機能
bHLH Transcription Factors in Cereal Crops: Diverse Functions in Regulating Growth, Development and Stress Responses
宋 宋 1、張 南南 1、范 暁偉 1,2,*、王 冠峰 3,4,*
1 College of Life Sciences, Henan Agricultural University, Zhengzhou 450002, China
2 Postdoctoral Station of Crop Science, Henan Agricultural University, Zhengzhou 450002, China
3 College of Agronomy and Biotechnology, China Agricultural University, Beijing 100193, China
4 Sanya Institute of China Agricultural University, Sanya 572024, China
* Correspondence: fanxw@henau.edu.cn (X.F.); gfwang@cau.edu.cn (G.W.)
Int. J. Mol. Sci. 2025, 26, 9915 https://doi.org/10.3390/ijms26209915
(要旨)
基本ヘリックス-ループ-ヘリックス(bHLH)転写因子は、イネ、トウモロコシ、小麦などの穀物作物において最大の転写調節因子ファミリーの一つである。これらは主要な農学的形質や必須生理過程の調節において重要かつ多様な役割を果たす。本総説は、三大穀物における機能的に特徴づけられたbHLH遺伝子を体系的に統合し、成長、発生、ストレス応答におけるそれらの役割について比較的な視点を提供する。我々は既知の機能を包括的にまとめ、穀粒サイズ制御のTaPGS1、葉角制御のイネILIサブファミリー、種子休眠制御のOsbHLH004、葯発達制御のトウモロコシ「Ms23-Ms32-bHLH122-bHLH51」カスケードなど特異的制御因子を強調する。鉄恒常性(例:IRO2)や干ばつ・低温・塩害・病原体応答における保存的機能と種特異的機能についても詳述する。さらに、E-box/G-boxシス要素への特異的結合、タンパク質二量体化、ホルモンシグナル経路との統合など、基盤となる分子メカニズムについても論じる。本総説は、現在の知見を統合することで、穀物の収量・品質・ストレス耐性向上を目指す分子育種プログラムにおけるbHLH転写因子の戦略的潜在性を強調する、包括的かつ最新の参考資料として機能する。
(以下、本文のうち、鉄栄養関連の部分だけを訳します)
3.5. 鉄恒常性
鉄は光合成、呼吸、窒素代謝など多くの重要な生理過程において決定的な役割を果たす。地球の地殻には豊富に存在するが、好気環境やアルカリ性土壌では生物学的利用能が著しく制限され、植物の鉄欠乏ストレスを頻繁に引き起こす。したがって穀物作物における鉄恒常性の維持は、収量と品質を確保する上で極めて重要である。植物は鉄を獲得するために主に二つの戦略を採用する:戦略Iと戦略II。戦略Iは主に双子葉植物およびイネ科以外の単子葉植物に見られる。植物が鉄欠乏シグナルを感知すると、根の細胞膜上の鉄還元酵素がFe(III)をFe(II)に還元し、これが鉄輸送体によって細胞内に輸送される。戦略IIはイネ科植物に特有である。鉄欠乏条件下で、植物はムギネ酸などの植物性シデロフォアを合成・分泌し、土壌中のFe(III)を効率的にキレート化してFe(III)–植物性シデロフォア複合体を形成する。これらの複合体は特定の輸送体によって根細胞内に輸送される。穀類作物では、複数のbHLH転写因子が複雑かつ精密な分子ネットワークを形成し、戦略IIを介した鉄恒常性を調節する。中核メカニズムには活性化、補助的調節、負のフィードバック平衡が含まれ、種間での保存性と特異性の両方を示す。
イネでは、OsIRO2(OsbHLH056)が鉄獲得の中核活性化因子として機能する。鉄欠乏条件下では、本因子が強く誘導され、植物性鉄輸送体合成関連遺伝子(例:OsNAS1/2、OsNAAT1、OsDMAS1)および輸送関連遺伝子(例:OsYSL15、TOM1)を活性化する。OsFIT(OsbHLH156)は物理的相互作用を介してOsIRO2の細胞内局在を調節する。通常条件下では、OsIRO2は主に細胞質に分布する。一方、鉄欠乏下では、OsFITがOsIRO2とヘテロ二量体を形成し、その核移行を促進することで下流遺伝子を活性化する。OsFITのノックアウトは、鉄欠乏下のイネにおいて葉の黄化と鉄含有量の減少を引き起こし、戦略II関連遺伝子(例:OsNAS2およびOsYSL15)の誘導発現を阻害する。これは、OsFITがOsIRO2機能に必須の補因子であり、両者が戦略IIの中核活性化モジュールを構成することを示唆している。一方、OsYSL15のノックアウトは、鉄欠乏下で葉の黄化を引き起こすものの、OsNAS2の発現には影響を与えない。OsIRO2の活性化作用とは対照的に、OsIRO3(同じくbHLH遺伝子)は負の制御によって鉄恒常性を維持する。OsIRO3の発現は鉄欠乏下で上昇し、過剰な鉄取り込みによる酸化損傷を防ぐため、OsIRO2やOsNAS3、OsYSL15などの下流遺伝子の発現を抑制する。その調節機構は、C末端のEARモチーフ(LxLxL)に依存している可能性があり、このモチーフがTOPLESSファミリー抑制因子をリクルートし、標的遺伝子の転写を阻害する。OsIRO3のノックアウトはイネにおいて過剰な鉄蓄積と葉の壊死を引き起こし、その負の調節役割を確認している。
イネの鉄恒常性には、複数の他のbHLH因子が関与している。これには正の調節因子であるOsbHLH060(OsPRI1)、sbHLH058(OsPRI2)、OsbHLH059 (OsPRI3)、OsbHLH133、および負の調節因子であるOsbHLH061が含まれる。OsPRI1、OsPRI2、OsPRI3は鉄欠乏下で誘導され、OsIRO2およびOsIRO3を活性化することで鉄吸収効率を高める。
OsbHLH061はPRI1と相互作用して複合体を形成し、PLESS/TOPLESSRELATED抑制因子(TPL/TPR)抑制因子を動員し、下流遺伝子 OsIRO2 および OsIRO3 を抑制する。この機構は鉄の長距離輸送を負に制御し、イネにおける鉄の均衡的な分布を確保する。別の遺伝子 OsbHLH133 は、おそらく根から茎への鉄の転送を負に制御することで、根と茎間の鉄の分配を調節する。トウモロコシと小麦における鉄恒常性の分子機構に関する研究は比較的限定的である(表4)。イネと同様に、トウモロコシのbHLH転写因子ペアであるZmFITとZmIRO2は相互作用してヘテロ二量体を形成し、植物性シデロフォアの合成と輸送に関わる遺伝子を活性化する。しかしながら、具体的な調節の詳細とネットワークは依然として不明である。小麦のトランスクリプトーム解析では、鉄欠乏が複数のbHLH遺伝子(例えば、イネのOsIRO2と相同性を持つTaIRO2など)の発現を誘導することが示されており、イネとの調節枠組みの保存可能性を示唆している。全体として、穀物作物におけるbHLH転写因子は「活性化-補助-抑制」の階層的ネットワークを通じて鉄恒常性を維持しており、鉄欠乏耐性作物の育種や栄養品質向上に向けた重要な標的を提供している。
5. 結論と展望
穀物作物における大規模かつ機能的に多様な転写調節因子ファミリーとして、bHLH転写因子は保存されたbHLHドメインを介してDNA結合とタンパク質相互作用を媒介し、成長、発生、代謝バランス、ストレス応答において精巧な調節ネットワークを構築する(図1)。成長・発生の観点では、穀粒形成、植物形態形成、葯発達から種子休眠に至るまで、bHLHファミリーはE-box/G-boxなどのシス要素への結合、ヘテロ二量体の形成、ホルモンシグナルへの応答を通じて、作物の主要な農学的形質を精密に制御する。ストレス適応においては、これらのbHLH転写因子の機能は「バランス」特性を示し、ABA、JA、SAなどのシグナルを統合することで、浸透圧調節、活性酸素種(ROS)消去、ストレス応答性遺伝子発現を調節し、最終的に作物の干ばつ、低温、塩ストレス、病害に対する抵抗性を高める。本総説では、イネ・トウモロコシ・小麦という三大穀物におけるbHLH調節因子の現状知見を統合した。
イネはモデル穀物として、葯発生における「UDT1-TIP2-TDR/EAT1」モジュールや種子休眠における「SD6/ICE2-OsbHLH048」経路など、明確に定義された制御カスケードを通じて最も包括的な知見を提供する。一方、トウモロコシや小麦はさらに大規模なbHLHファミリーを有するものの、特に小麦において、植物形態形成や生物的ストレス応答などのプロセスにおける構成員の機能解明は遅れている。この格差は重大な知識の空白を浮き彫りにする。トウモロコシと小麦は広範な栽培と世界食糧安全保障における中核的役割から、分子レベルの深い理解が求められるため、この空白を埋めることは特に重要である。
これまでの顕著な進展にもかかわらず、穀物改良におけるbHLH因子の潜在能力を完全に実現するには、いくつかの重要な科学的課題がさらなる探求を必要としている:
bHLHファミリーにおける機能的冗長性の分子基盤。bHLHファミリーは多数の構成員を有し、同一サブファミリー内や異なるサブファミリー間で機能的冗長性が頻繁に生じる[3]ため、その機能研究は複雑化している。機能的冗長性と機能特異的制御の分子基盤を理解することは、現在の研究における重要な課題である。
今後の研究では、遺伝学的手法、分子生物学的手法、バイオインフォマティクス的手法を組み合わせ、bHLHファミリーメンバー間の相互作用ネットワークと、機能的冗長性と特異的調節のバランスを体系的に解析する必要がある。
bHLHタンパク質の精巧な調節ネットワーク。これまでbHLH転写因子の機能解析は、限られたモデル遺伝子と主要な生理過程に焦点が当てられてきた一方で、ファミリー全体の機能に関する未知の領域は依然として多い。特にトウモロコシや小麦では、植物形態形成やストレス応答などの側面におけるbHLH転写因子の研究は比較的遅れている。先に指摘した通り、この知識のギャップを埋めることは喫緊の課題である。同時に、既知の機能を持つbHLH転写因子の調節ネットワークの精密なメカニズム、例えば他のシグナル伝達経路とのクロストークや、翻訳後修飾が機能を調節する具体的な分子機構などについては、依然として深い調査が必要である。これらの疑問に取り組むには、先進的な技術的手法と統合したより詳細な研究が不可欠である。
bHLH遺伝子の育種応用戦略。穀物作物におけるbHLH転写因子の機能と調節機構に関する詳細な研究は、分子育種技術を用いた作物形質の改良に向けた豊富な遺伝子資源と理論的指針を提供する。CRISPR/Cas9などの遺伝子編集技術を用いることで、主要なbHLH遺伝子を改変することで、作物の収量、品質、ストレス耐性を向上させることが可能である。例えば、イネSD6およびその小麦相同遺伝子を編集することで、収穫前発芽を抑制できる[77]。さらに、bHLH遺伝子の自然変異に基づく機能的分子マーカーの開発が可能である。例えば、小麦TabHLH27-A1のプロモーター領域における自然変異は、干ばつ耐性、収量、水利用効率と関連しており、マーカー支援選抜育種のための分子マーカーとして利用可能である。トウモロコシZmbHLH124の耐乾性対立遺伝子も、耐乾性遺伝資源を選別するための分子マーカー開発に利用可能である。これらの機能マーカーの活用は、穀物作物の重要形質におけるピラミッド育種プロセスを加速し、育種効率を向上させ得る。
以下は図1の説明.
穀物作物におけるbHLH転写因子の分子機構と多様な機能の概要図。bHLH転写因子は、E-boxモチーフ(CANNTG)へのDNA結合を促進する塩基性領域と、タンパク質二量体を媒介するヘリックス-ループ-ヘリックス(HLH)ドメインという2つの保存領域を特徴とする。bHLHタンパク質は、典型/非典型bHLHタンパク質や異なるタイプの転写因子とホモ二量体またはヘテロ二量体を形成し、その調節スペクトルと特異性を大幅に拡大する。その活性は、ABA、BR、JAなどのホルモンシグナル伝達経路と統合されることが多い。マスターレギュレーターとして機能するbHLH転写因子は、穀物作物のライフサイクル全体にわたり、穀粒発達、植物形態形成、葯・花粉形成、鉄恒常性、非生物的ストレス(干ばつ、低温、塩害など)および生物的ストレス応答を含む多様な重要な農学的形質を統括する。各プロセスにおいて、イネ、トウモロコシ、小麦から機能的に特徴づけられた代表的遺伝子を重点的に取り上げる。
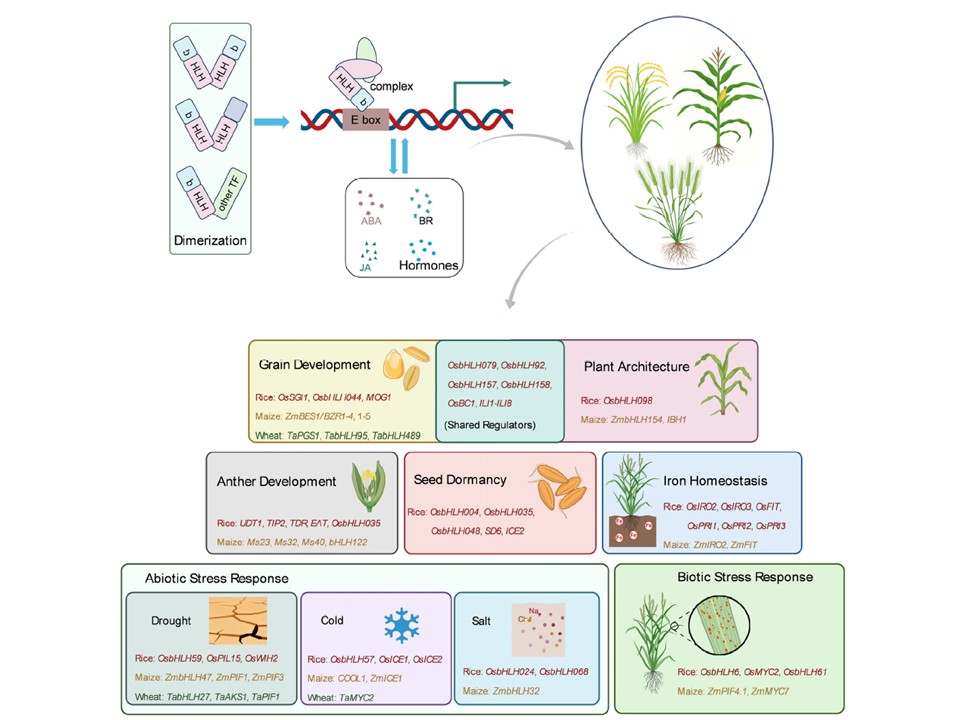
図1.穀物作物におけるbHLH転写因子の分子機構と多様な機能の概要図
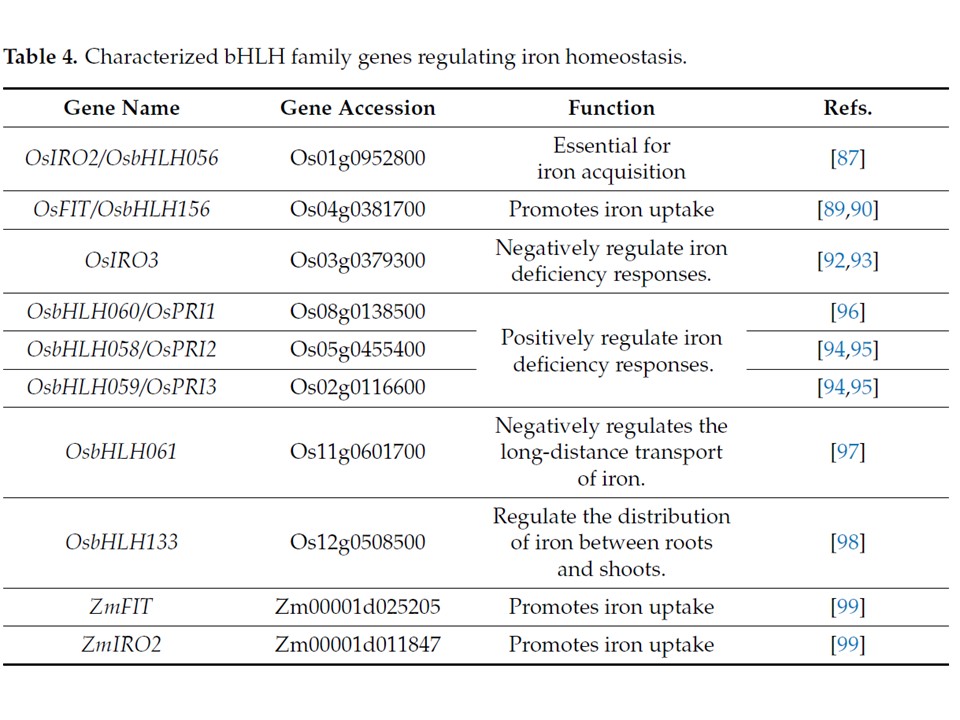
表4.鉄恒常性を調節する特徴付けられたbHLHファミリー遺伝子